
Гаплогруппа R1b

Европейские языки зародились на Армянском нагорье

Совершенно новое исследование, опубликованное в последнем выпуске журнала Science, раскрывает происхождение индоевропейской языковой семьи, расположенной на древнем Армянском нагорье. Так называемая теория анатолийской прародины, впервые предложенная в конце 1980-х годов профессором Колином Ренфрю (ныне лорд Ренфрю), получила постепенное признание, но оставалась спорной, пока новый метод изучения смещения языка не был представлен доктором Квентином Аткинсоном из Оклендского университета в Новой Зеландии. Используя новые научные методы, полученные из эволюционной биологии, доктор Аткинсон и его команда объявляют о том, что они раскрыли тайну происхождения индоевропейской языковой семьи. Доктор Аткинсон и его команда создали базу данных, содержащую 207 родственных слов, присутствующих в 103 индоевропейских языках, включая 20 древних языков, таких как латынь и греческий. Используя филогенетический анализ, они смогли реконструировать эволюционное родство этих современных и древних языков — чем больше родственных слов, тем более похожи языки и тем ближе они группируются на древе. Индоевропейская семья является одной из крупнейших семей — более 400 языков, на которых говорят по крайней мере в 60 странах. Результат, как они объявили в четверговом выпуске журнала Science, заключается в том, что «мы нашли решающее подтверждение анатолийского происхождения, а не степного». И время, и корень древа индоевропейских языков «соответствуют сельскохозяйственной экспансии из Анатолии, начавшейся 8000–9500 лет назад», сообщают они.
Как язык, родившийся на Армянском нагорье, армянский также принадлежит к индоевропейской семье (как видно на диаграмме ниже). Мы также можем наблюдать из древа, армянский язык, по-видимому, является одним из древнейших сохранившихся языков, которые используются и сегодня.

Эти фонды получили освещение в ведущих новостных агентствах по всему миру.
Гипотеза Армянского нагорья становится все более правдоподобной

Идея о том, что большинство европейских языков родственны и, возможно, происходят от общего предкового языка, известного как протоиндоевропейский (ПИЕ), была хорошо принята учеными различных дисциплин. Так называемая индоевропейская языковая семья включает большинство европейских, а также некоторые языки Плодородного полумесяца, Кавказа, Западной и Южной Азии. С более чем 400 языками (включая диалекты) она является крупнейшей языковой семьей в мире и на ней говорят почти 3 миллиарда носителей языка по всему миру. Некоторые считают армянский язык одним из древнейших сохранившихся членов этой семьи, по некоторым оценкам, он существовал еще 5000 лет до нашей эры. Однако предполагаемая родина носителей протоиндоевропейского языка стала предметом многочисленных споров. Существует четыре конкурирующие гипотезы, которые помещают происхождение индоевропейской языковой семьи либо в Понтийскую степь, Анатолию, Балканы или Армянское нагорье. Традиционная точка зрения помещает родину в Понтийские степи около 6000 лет назад. Альтернативная гипотеза утверждает, что языки распространились из Анатолии с расширением земледелия 8000–9500 лет назад. Еще одна гипотеза помещает родину протоиндоевропейского языка на Балканы, откуда индоевропейские земледельцы предположительно распространили язык по всей Европе. И последняя гипотеза называет Армянское нагорье наиболее вероятным кандидатом на праиндоевропейскую родину.
Долгожданное исследование древней ДНК, недавно опубликованное на сайте препринтов bioRxiv, пролило больше света на эту проблему с довольно интересными и, возможно, более загадочными результатами относительно роли ранних армян в распространении индоевропейского языка и генов. Работа была проделана большой группой под руководством генетиков Дэвида Райха и Иосифа Лазаридиса из Гарвардской медицинской школы в Бостоне и Вольфганга Хаака из Университета Аделаиды в Австралии. В исследовании использовались самые современные методы ДНК для анализа древней ДНК 69 европейцев, живших между 8000 и 3000 лет назад, чтобы генетически отследить перемещения древнего населения.



J2- Малая Армения Киликия и Армянское междуречье – Ряд современных генетиков считает, что J2 возник на Армянском нагорье. – Родина тех, у кого есть ген J2 – Малая Армения, Киликия и Армянское междуречье. J2 происходит от исходной oplogrupa R1b. Родина которой – Великая Армения!
R1b – Великая Армения – Ряд современных генетиков считает, что R1b возник на Армянском нагорье.
История исследований
1) Институт молекулярной и клеточной биологии Отдел эволюционной биологии Урмас Русталу Мн. RVA-Лиз Лугвали Проф. д-р Ричард Виллемс Тарту 2004 «В нашем исследовании предки армян были прослежены до разных частей Армении, Грузии, Азербайджана и Турции, что указывает на то, что историческая Армения была гораздо большей территорией, чем нынешняя Республика Армения».
2) Баноеи, Чалештори, Саньяти, Хаушман, Маджидзаде, Солтани и Голиапур (2007) Изменчивость аллелей и генотипов DAT1 VNTR среди старых этнических групп в Месопотамии до региона Окс. «Армяне — это нация и этническая группа, происходящие с Армянского нагорья, где наблюдается большая концентрация этого сообщества, особенно в Армении». 3) (Мовсесян и Кочар 2000) «Сходство черепов современных армян и жителей Армении 1600 – 700 гг. до н.э. свидетельствует о преемственности генетической связи с древним населением»
4) «Армяне – это отдельная этническая группа, которая произошла от неолитических племен Армянского нагорья» Литвинов С*, Кутуев А, Юнусбаев В, Р. Хусаинова, Валиев Р, Хуснутдинова Э.
5) 40% армянских генов относятся к эпохе палеолита. Левон Епископосян
6) Армяне относятся к 13 различным генетическим группам, возраст которых исчисляется десятками тысяч лет, и при этом в ДНК армян за последние 4000 лет нет следов захватчиков, что делает их «однородными во всем их многообразии».
В результате тестирования более 300 человек было установлено, что армянские ветви ДНК являются основой многих европейских ветвей.
Разные народы в разное время использовали разные названия для Армении и армян. Шумеры около 2800 г. до н. э. называли Армению Араттой, а аккадцы, пришедшие на смену шумерам во второй половине третьего тысячелетия до н. э., называли Армению Армани или Арманум.
Хетты, появившиеся во втором тысячелетии до н. э., использовали для обозначения Армении название Хайаса, а ассирийцы, возникшие во второй половине второго тысячелетия до н. э., использовали название Уруатри или Урарту (Арарат в Библии).
С исчезновением с исторической арены народов, называвших Армению и армян этими именами, исчезло и использование этих названий. Но Армения и армяне, которые всегда находили в себе силы и возможность обновляться на протяжении всего этого времени, продолжали и продолжают жить.
Аратта считается первым армянским государством, упомянутым в древних текстах. Это записи древнего шумерского эпоса о Гильгамеше. Считается, что этим текстам около 4800 лет, но это не значит, что государство Аратта не существовало до этого.
Квентин Аткинсон и Рассел Грей доказали, что армянский язык уже отделился от общего праиндоевропейского языка на Армянском нагорье, прародине индоевропейцев, около 8500 лет назад.
Следует помнить, что наши предки построили храмовый комплекс Портасар около 12 000 лет назад, что свидетельствует о высоком уровне социальной организации, который, безусловно, был необходим для выполнения такой грандиозной работы по меркам того времени.
То, что армяне являются коренными жителями Армянского нагорья, уже хорошо известно из генетических исследований. Например, исследование Хабера и др. (2015) обнаружило непрерывный армянский ген на Армянском нагорье в течение 4000 лет. Результаты теста гласят:
«Наши тесты показывают, что армяне не имели значительного смешения с другими популяциями в своей недавней истории и, следовательно, были генетически изолированы с позднего бронзового века».
«Положение армян в глобальном генетическом разнообразии уникально и, по-видимому, отражает географическое положение Анатолии (читай Армянского нагорья, как далее). Принятие армянами отличительной культуры на раннем этапе их истории привело к их генетической изоляции от своего окружения».
Такой же вывод сделали Хелленталь и др. (2014) в своем Генетическом атласе истории смешения людей, опубликованном в журнале Science. Из-за этих результатов некоторые ученые называют современных армян «живым ископаемым».
Другие исследования, изучающие древнюю ДНК, собранную из захоронений, обнаружили генетическое сходство между современными армянами и древними жителями Армянского нагорья.
Аллентофт и др. (2015), например, наблюдали генетическое сходство между людьми бронзового века (ок. 3500 лет назад) и современными армянами, а Лазаридис и др. (2016) показали сходство между людьми медного века (ок. 6000 лет назад) и людьми бронзового века (ок. 3500 лет назад), раскопанными в Армении.
Вооружившись этими выводами, Левон Епископосян, руководитель лаборатории Института молекулярного биоразнообразия Национальной академии наук, заявил на пресс-конференции, что:
«Результаты генетических исследований показали, что образцы ДНК людей бронзового века, найденные на территории Армении, имеют генетическое сходство с генетикой современных людей, живущих в Армении сегодня», «Современные армяне являются прямыми потомками людей, которые жили на территории Армении 5000 лет назад».
Похожие заявления сделал известный блогер-генетик Диенекес, где он подтверждает генетическую преемственность армян, но задается вопросом, распространяется ли эта преемственность за пределы бронзового века:
«Когда речь идет о Кавказе/Ближнем Востоке, при более близком рассмотрении становится ясно, что армяне бронзового века почти неотличимы от современных армян. Сохранилась ли генетическая преемственность армян в более ранние периоды?»
Новые данные показывают, что эта преемственность на самом деле распространяется за пределы бронзового века (на основе тестирования митохондриальной ДНК), уходя на 7811 лет назад.
Митохондрии передаются от матерей к их детям. Таким образом, изучение митохондриальных геномов позволяет ученым проследить уникальную историю женской линии с течением времени.
Это новое исследование, изучающее древнюю материнскую ДНК из скелетных останков, раскопанных в Армении и Арцахе, подтвердило сильное сходство с ДНК современных армян.
Исследование «Восемь тысячелетий генетической преемственности по материнской линии на Южном Кавказе», опубликованное в журнале Current Biology, изучило 52 древних генома из останков скелетов, раскопанных в Армении и Арцахе. Калиброванные радиоуглеродные даты древних образцов составили от 300 до 7811 лет до н. э.
«Мы проанализировали множество древних и современных митохондриальных геномов в некоторых частях Южного Кавказа и обнаружили генетическую преемственность в течение как минимум 8000 лет», — сказали Ашот Маргарян и Мортен Э. Аллентофт из Центра географии в Музее естественной истории Дании.
«Другими словами, мы не смогли обнаружить никаких изменений в женском генофонде за этот очень долгий период времени. Это очень интересно, потому что этот регион пережил много культурных сдвигов за тот же период, но эти изменения, похоже, не оказали генетического воздействия — по крайней мере, на женское население».
The researchers were interested in studying this part of the world because of its position as a cultural crossroads since ancient times. It is also known as an important area for the potential origin and spread of Indo-European languages.
В исследовании говорится:
Этот результат предполагает, что за последние 7800 лет не произошло никаких серьезных генетических сдвигов в генофонде мтДНК в Армянском нагорье. Мы обнаружили, что наименьшее генетическое расстояние в этом наборе данных находится между современными армянами и древними людьми, что также отражено как в сетевом анализе, так и в дискриминантном анализе главных компонент.
Армяне из разных регионов, включая Эрзурум, Арарат и Арцах, показали самую прямую связь с древними жителями Армянского нагорья.
Очевидно, что современные армянские группы и древняя группа имеют очевидные сходства.
Кроме того, в статье отмечено заметное сокращение женского населения около 25 000 лет назад во время последнего ледникового периода (ПЛП), за которым последовал быстрый (примерно в 10 раз) рост населения около 10 000 лет назад (см. рисунок), что показывает быстрый рост населения в период неолита, когда люди впервые открыли для себя сельское хозяйство.
Эти результаты имеют важные последствия для мирового научного сообщества. Похоже, что за последние восемь тысячелетий в генофонде армян не произошло никаких серьезных генетических изменений, несмотря на многочисленные хорошо документированные культурные изменения в регионе.
Археологически и исторически задокументированные миграции центральноазиатских групп (например, турок и монголов) в Армянское нагорье, по-видимому, не внесли существенного вклада в материнский генофонд армян.
Как географические (горные регионы), так и культурные факторы могли служить барьерами для генетических контактов между армянами и мусульманскими захватчиками в XI-XIV веках н. э.
Ген армянских женщин не сильно изменился за 8000 лет
Первые данные генома из древних египетских мумий
Датские, армянские, российские и британские ученые обнаружили, что женское население Армении и Арцаха практически не изменилось с генетической точки зрения за последние восемь тысяч лет.
Соответствующее исследование было опубликовано в журнале Current Biology, а Cell Press кратко о нем сообщил.
«Мы не смогли обнаружить никаких изменений в женском генофонде за очень длительный период времени. Это крайне интересно, поскольку регион пережил множество культурных сдвигов за тот же период времени, но эти изменения, вероятно, не связаны с генетическим влиянием, по крайней мере, на женскую популяцию», — заявили соавторы исследования Ашот Маргарян и Мортен Аллентофт.
К таким выводам специалисты пришли после анализа полных митохондриальных геномов 52 древних скелетов из Армении и Нагорного Карабаха, возраст самых старых образцов составляет 7,8 тыс. лет. Ученые сравнили эти данные с информацией о митохондриальных геномах 206 современных армян, а также с ранее опубликованными материалами о более чем 480 людях из семи популяций, граничащих с Арцахом.
Анализ показал, что численность населения на территории современных Армении и Арцаха быстро увеличивалась после последнего ледникового максимума (около 18 тыс. лет назад), однако миграции в регионе, произошедшие около двух-трех тысяч лет назад, практически не затронули ее женскую часть. В дальнейшем ученые планируют провести генетический анализ образцов, взятых с восточных территорий Азербайджана, а также Грузии.
То, что армяне являются коренными жителями Армянского нагорья, уже хорошо известно из генетических исследований. Например, исследование Хабера и др. (2015) обнаружило непрерывный армянский ген в Армянском нагорье на протяжении 4000 лет. Результаты теста гласят:
«Наши тесты показывают, что армяне не имели значительного смешения с другими популяциями в своей недавней истории и, следовательно, были генетически изолированы с конца бронзового века».
«Положение армян в глобальном генетическом разнообразии уникально и, по-видимому, отражает географическое положение Анатолии (читай Армянского нагорья, как в следующем тексте). Принятие армянами отличительной культуры на раннем этапе их истории привело к их генетической изоляции от своего окружения».
Тот же вывод был сделан Хелленталем и др. (2014) в их Генетическом атласе истории смешения людей, опубликованном в журнале Science. Из-за этих результатов некоторые ученые называют современных армян «живым ископаемым».
Другие исследования, изучающие древнюю ДНК, собранную из захоронений, обнаружили генетическое сходство между современными армянами и древними жителями Армянского нагорья.
Например, Аллентофт и др. (2015) наблюдали генетическое сходство между людьми бронзового века (около 3500 лет назад) и современными армянами, а Лазаридис и др. (2016) показали сходство между людьми медного века (около 6000 лет до н. э.) и людьми бронзового века (около 3500 лет до н. э.), раскопанными в Армении.
Подготовив такие выводы, заведующий лабораторией Института молекулярного биоразнообразия Национальной академии наук Левон Епископосян заявил на пресс-конференции, что:
Результаты генетических исследований показали, что образцы ДНК людей бронзового века, найденные в Армении, имеют генетическое сходство с генетикой современных людей, живущих в Армении сегодня», «Современные армяне являются прямыми потомками людей, которые жили в Армении 5000 лет назад».
Подобные заявления сделал известный блогер-генетик Диенекес, где он подтверждает генетическую преемственность армян, но задается вопросом, распространяется ли эта преемственность за пределы бронзового века:
«Когда речь идет о Кавказе/Ближнем Востоке, при более близком рассмотрении становится ясно, что армяне бронзового века почти неотличимы от современных армян. Продолжается ли генетическая преемственность армян в более ранние периоды?”
Новые данные показывают, что эта преемственность действительно простирается за пределы бронзового века (на основе тестов митохондриальной ДНК), уходя на 7811 лет назад.
Митохондрии передаются от матерей к детям. Таким образом, изучение митохондриальных геномов позволяет ученым проследить уникальную историю женской линии с течением времени.
Это новое исследование, изучающее древнюю материнскую ДНК из скелетных останков, раскопанных в Армении и Арцахе, подтвердило сильное сходство с ДНК современных армян.
Исследование «Восемь тысячелетий матрилинейной генетической преемственности на Южном Кавказе», опубликованное в журнале Current Biology, изучило 52 древних генома из скелетных останков, раскопанных в Армении и Арцахе. Калиброванные радиоуглеродные даты древних образцов составили от 300 до 7811 лет до н. э.
«Мы проанализировали множество древних и современных митохондриальных геномов в некоторых частях Южного Кавказа и обнаружили генетическую преемственность на протяжении как минимум 8000 лет», — говорят Ашот Маргарян и Мортен Э. Аллентофт из Центра географии в Музее естественной истории Дании.
«Другими словами, мы не смогли обнаружить никаких изменений в женском генофонде за этот очень долгий период времени. Это очень интересно, потому что этот регион пережил много культурных сдвигов за тот же период, но эти изменения, похоже, не оказали генетического воздействия — по крайней мере, на женское население».
Исследователи были заинтересованы в изучении этой части мира из-за ее положения как культурного перекрестка с древних времен. Она также известна как важная область для потенциального происхождения и распространения индоевропейских языков.
В исследовании говорится:
Этот результат предполагает, что за последние 7800 лет не произошло никаких серьезных генетических сдвигов в генофонде мтДНК в Армянском нагорье. Мы обнаружили, что наименьшее генетическое расстояние в этом наборе данных находится между современными армянами и древними людьми, что также отражено как в сетевом анализе, так и в дискриминантном анализе главных компонент.
Армяне из разных регионов, включая Эрзурум, Арарат и Арцах, показали самую прямую связь с древними жителями Армянского нагорья.
Очевидно, что современные армянские группы и древняя группа имеют очевидные сходства.
Кроме того, в статье отмечено заметное сокращение женского населения около 25 000 лет назад во время последнего ледникового периода (ПЛП), за которым последовал быстрый (примерно в 10 раз) рост населения около 10 000 лет назад (см. рисунок), что показывает быстрый рост населения в период неолита, когда люди впервые открыли для себя сельское хозяйство.
Эти результаты имеют важные последствия для мирового научного сообщества. Похоже, что за последние восемь тысячелетий в генофонде армян не произошло никаких серьезных генетических изменений, несмотря на многочисленные хорошо документированные культурные изменения в регионе.
Археологически и исторически задокументированные миграции центральноазиатских групп (например, турок и монголов) в Армянское нагорье, по-видимому, не внесли существенного вклада в армянский материнский генофонд.
Как географические (горные регионы), так и культурные факторы (индоевропейские христиане и тюркоязычные мусульмане) могли служить барьерами для генетических контактов между армянами и мусульманскими захватчиками в XI-XIV вв. н. э.
Ген армянских женщин практически не изменился за 8000 лет
Датские, армянские, российские и британские ученые обнаружили, что женское население Армении и Арцаха практически не изменилось с генетической точки зрения за последние восемь тысяч лет.
Соответствующее исследование было опубликовано в журнале Current Biology, а Cell Press кратко о нем сообщил.
«Мы не смогли обнаружить никаких изменений в женском генофонде за очень длительный период времени. Это чрезвычайно интересно, поскольку регион пережил множество культурных сдвигов за тот же период времени, но эти изменения, скорее всего, не связаны с генетическим влиянием, по крайней мере, на женское население», — заявили соавторы исследования Ашот Маргарян и Мортен Аллентофт.
К таким выводам специалисты пришли после анализа полных митохондриальных геномов 52 древних скелетов из Армении и Нагорного Карабаха, самые старые образцы которых имеют возраст 7,8 тысяч лет. Ученые сопоставили эти данные с информацией о митохондриальных геномах 206 современных армян, а также ранее опубликованными материалами по более чем 480 людям из семи популяций, граничащих с Арцахом.
Анализ показал, что численность населения на территории современной Армении и Арцаха быстро увеличивалась после последнего ледникового максимума (около 18 тысяч лет назад), однако миграции в регионе, имевшие место около двух-трех тысяч лет назад, практически не затронули его женскую часть. В дальнейшем ученые планируют провести генетический анализ образцов, взятых с восточных территорий Азербайджана, а также Грузии.
В этом контексте «вырисовывается» весьма интересная гипотеза, которая может пролить свет на все это и помочь собрать мозаику в единую картину.
Я имею в виду гиксосов. Для того, чтобы…
1-я официальная теория о «гиксосах» заключается в том, что в 1700-1650 годах до н. э. армянские племена захватили власть в Древнем Египте. Они основали свою столицу (Аварис) и правили Египтом более 100 лет, за это время, по разным данным, сменилось 6-7 «гиксийских» царей.
Но в этой теории много ляпов и подмен, а также слишком много того, что указывает на нечто совсем иное, нежели семитские кочевники-скотоводы. И даже не один наводящий на размышления факт, а целый набор, причем взаимосвязанных.
Сам факт. Как племя кочевников-скотоводов могло завоевать ослабленную, но все еще великую цивилизацию? И не просто завоевать, а править ею более 100 лет? – Никак нет
2-я официальная теория
Вторая официальная теория и самая распространенная теория о «гиксосах» заключается в том, что в 1700-1650 гг. до н. э. армянские племена — оставшаяся часть армянского народа — захватили власть в Древнем Египте. Они основали свою столицу (Аварис) и правили Египтом более 100 лет, за это время, по разным данным, сменилось 6-7 «гиксийских» царей.
Название. Название «гиксосы» (точнее гиксос/хайксос) происходит из греческих источников.
В попытке связать его с египетским лексиконом «ученые» адаптировали к данному случаю египетское «hqa xaswt» (hekau khasut) — правители чужой страны, или — правители чужих стран. При этом они не удосуживаются объяснить, как греки могли так исказить это название. Поэтому остановимся на греческом оригинале, а не на «домыслах» о египетском значении.
Окончание «сос» — явно греческое дополнение, которое было присоединено в результате усилий эллинизированных Манефона, Иосифа Флавия и Евсевия Кесарийского (основных источников по Айксосу). Когда мы убираем окончание «сос», остается изначальная форма слова — «айк». А теперь подвох.
Самоназвание армян во все времена было «хай» (армянин), а в древнеармянском окончание -к давало множественное число «айк» (армяне). Связано это с легендой об отце-прародителе всех армян по имени Айк.
Завоевать Египет с небольшой армией и удерживать его более века могли только пришельцы с гораздо более высоким уровнем технологического развития, и совершенно очевидно, что семитские кочевники-скотоводы к этой категории относиться не могли. Этот пункт (как и первый) подтверждается археологическими находками: 3. Достоверно известно, что гиксосы прибыли на колесницах, это подтверждают как египетские изображения, так и сохранившиеся образцы колесниц гиксосов в пирамидах. Ни у египтян до этого, ни у семитских кочевников в то время колесниц не было вообще. А в Армении, в Айасе, использовали колесницы. 4. Колесницы были сделаны из дерева, которое в то время росло только на территории Армянского нагорья (Армении). Сомневаюсь, что семиты тащились бы в наше нагорье за дровами… 5. Более того. В конструкции этих самых колесниц обнаружены железные детали, что говорит о том, что гиксосы владели развитыми технологиями металлургии (обработки железа). Их оружие также было железным, о чем свидетельствует, например, череп последнего фараона Среднего царства, прорубленный через бронзовый шлем. До гиксосов египтяне владели только бронзой, не говоря уже о семитских скотоводах. 6. Идем дальше. Где в то время была развита металлургия? Ни в степях Средней Азии, ни на территории современной Персии железо еще не обрабатывалось. А обрабатывали его в то время на Армянском нагорье. Об этом свидетельствуют археологические раскопки, найден ряд металлоплавильных центров (например, в Мецаморе), где металл выплавляли в больших масштабах уже в начале II тысячелетия. 7. и в подтверждение пункта 6 – именно Армянское нагорье, а не соседние регионы, богато железной рудой, которую, как доказали археологи, начали добывать уже в 3 тысячелетии. 8. Гиксосы говорили, что у них был свой царь (на их родине) и называли его Хианом. В армянских легендах есть сказания об армянском царе по прозвищу Хианали, “хианали” в переводе с армянского – красивый. 9. Они сначала назвали столицу Хайару, а затем переименовали ее в Аварис. Оба названия имеют армянскую трактовку. Хайару дословно переводится как “армянский мужчина”, это название можно связать с тем, что изначально город был военным лагерем, в котором жили только мужчины. Что касается Авариса, то тут в целом все понятно – дословный перевод “мой трофей”.
- Менее значимый и ясный факт, но тем не менее также известно, что гиксосы называли свои колесницы «варака», по-армянски глагол «варел» — водить, и употребляется в такой форме только тогда, когда речь идет об управлении каким-либо видом транспорта.
- По описаниям и найденным изображениям «гиксосы» очень напоминают армянскую расу — арменоидов. В частности, большие миндалевидные глаза — «фишка» армян.
- В пантеоне «айксосов» ведущее место занимали бог грома и его жена Анаит. Богиня Анаит — главная богиня армянского пантеона, а «бог грома» — армянский бог войны Вааг, считавшийся повелителем грома.
- Правящая династия «Айксов» в источниках именуется «хекаказут» (С. Галямов «Башкоры от Гильгамеша до Заратустры», Уфа 1999), что есть не что иное, как название одной из ветвей армянского царского рода того времени «айказуни» и на самом деле имеет форму «айказут» («зут» имеет уменьшительное значение в армянском языке и символизирует то, что «айказут» — «младшие братья» правящего дома «айказуни»).
- «Айксос» ввел 12-знаковый зодиак в Египте, и по одной из гипотез, он появился именно на Армянском нагорье, потому что только в фауне и культуре Армянского нагорья присутствует полный набор представителей 12 знаков — ни в одной другой стране КОМПЛЕКСА нет Овна, Козерога, Тельца, Льва (они были очень распространены на Армянском нагорье), а также Водолея (культ воды был чрезвычайно развит на Армянском нагорье), Стрельца (Бог армян того времени Айк имеет образ стрелка (лучника)). О степени развития астрологии у армян говорит и тот факт, что на территории Армянского нагорья было найдено несколько древних обсерваторий. Например, Караундж, которой более 7500 лет. (ссылка)
- Именно в период 1750-1650 гг. до н.э. (не раньше и не позже) у армян была прекрасная возможность для столь глубоких походов на юг, поскольку все соседние с юга страны, которые могли бы этому противостоять, были в то время крайне ослаблены или еще не достигли такого расцвета:
Сам Египет сильно ослаб и сошёл с арены международных противоборств. Сирийские города-государства после нашего нашествия хамураппи перестали из себя чего-то представлять, так же как и сама Вавилон во главе с хамураппи была только на чале своего возбуждения и не представляла еще опасности. Крайняя плита была Эламом, который возвысится только позже, то же можно сказать про Хетти. Ассирия была разделяема внутренне борьбой за власть и тоже была крайне слаба… Кстати, именно в последнем случае мое имеющее прямое доказательство – по данным клинописей именно в 1700-ом году в Асирии закончилась борьба за власть , что без сомнений можно объяснить завоевание арменоидами. Более того, по сведениям арабских источников (Масуди) армане победили ассирийского царя Арсиса (видимо имеется ввиду Адаси, который процарствовал меньше года и именно в 1700-ом), после чего 20 царей Ассирии в течении 273 лет подчинялись армянам и платили дань… Подитожим: Эти пункты не только объясняют то, как могла небольшая армия чужеземцев поработать великую цивилизацию более чем на век (благодаря технологическому превосходству), но и заставляет скептически относиться к теории о семитском происхождении Хайксосова и взглянуть по другому на вопрос об их происхождении. Ибо такое количество “совпадений” не может быть случаем К чему я вообще написал про Хайксосова и как это связано с гаплогруппами. Обратите внимание на период упоминаний о Хайксосах – примерно 3500-3700 лет назад. Это именно тот период, когда у армян появился второй гаплотип – R1b. Учитывая всё вышесказанное про Хайксосова, а так же полное совпадение по временам появления у армян гаплотипа Р1б можно предложить следующий сценарий. Возможно древние армяне (у которых еще была лишь группа J2) примерно 3500-4000 лет назад совершили несколько дальних военных походов с армянского нагорья. Как писалось выше, некоторые учения называют более точные даты 1750-1650 годов д.н.э. относительно походов на юг (Египет), о чем было сказано чуть выше. И тут встает вопрос! Если древние армяне, могли бы себе позволить подобные военные походы с армянского нагорья на юго-юго-запад, то им тем более ничего не мешало совершать подобные же походы на запад, через Босфор в Европу? Армяне дойдя до Европы – продолжительное время там жили (пару тройку сотен лет), ассимилировали какое-то местное население и впоследствии вернулись обратно, вдобавок к новым генам в себе привели с собой определенную часть европейцев. К тому же продолжительное проживание на совершенно другом ареале так же может способствовать мутациям Скорое всех древних армяне завершили поход в том числе и в этом направлении, впоследствия “принесли” с собой на армянское нагорье западноевропейский гаплотип Р1б. P.S. Вообще история армян достаточно прозрачна и неоспорима в основе вплоти до ~1000 лет д.н.э. А сценарии “до этого” – есть разные. 10000 и более пусть назат сформировалась наша нация . В поддержку аргументов есть, правда небольшого. Правда эти “осколки” порой собираются в очень четкие логические цепочки. Но к сожалению процесс глубокого познания затрудняет множество факторов. 1. Армяне воевали практически всю свою историю, наши противники зачастую не брезгали попытками массового истребления, уничтожения культурного наследия. Многое было утрачено в последствии войны, зависти соседей и жажды присвоения. Кстати, жада присвоения есть и посей день, например в той же Грузии она обрела второе дыхание, об этом можно почитать тут. Там документы, фото… вот один из огромного числа примеров “грузинизации” армянских памятников из этого материала: 3. Сейчас вести раскопки на Армянском Нагорье нереально, туркам это ой как не выгодно…они за последние 90 лет уничтожили большинство армейского культурного наследия в западной Армении, а тут еще и новое раскапывать… Пока там турки – ничего Армянского там найти будет нельзя, а если и будут находить случайно – то все следы принадлежности к Армянам будут уничтожиться, либо если это нереально – уничтожается сама находка.
Сам Египет был сильно ослаблен и сошёл с арены международных противостояний. Сирийские города-государства после нашествия Хамурапи перестали быть чем-либо, как и сам Вавилон под предводительством Хамурапи находился только в начале своего подъёма и ещё не представлял угрозы. Крайне слаб был Элам, который поднимется лишь позже, то же самое можно сказать и о Хеттии. Ассирия была раздроблена внутри борьбой за власть и также была крайне слаба… Кстати, именно в последнем случае у нас есть прямые доказательства — согласно клинописным данным, именно в 1700 году в Ассирии закончилась борьба за власть, что, несомненно, можно объяснить завоеванием арменоидами. Более того, согласно арабским источникам (Масуди), армяне победили ассирийского царя Арсиса (видимо, имеется в виду Адаси, правивший менее года, а именно в 1700 году), после чего 20 царей Ассирии на протяжении 273 лет подчинялись армянам и платили дань… Подводя итог: Эти моменты не только объясняют, как небольшая армия иноземцев могла более века порабощать великую цивилизацию (за счет технологического превосходства), но и заставляют нас скептически отнестись к теории семитского происхождения гиксосов и по-другому взглянуть на вопрос их происхождения. Потому что такое количество «совпадений» не может быть случайным. Зачем я вообще написал о гиксосах и как это связано с гаплогруппами? Обратите внимание на период упоминаний о гиксосах — примерно 3500-3700 лет назад. Это как раз тот период, когда у армян появился второй гаплотип — R1b. Учитывая все вышесказанное о гиксосах, а также полное совпадение по времени появления гаплотипа R1b у армян, можно предположить следующий сценарий. Возможно, древние армяне (у которых осталась только группа J2) совершили несколько длительных военных походов с Армянского нагорья примерно 3500-4000 лет назад. Как было написано выше, некоторые ученые называют более точные даты 1750-1650 гг. до н.э. относительно походов на юг (Египет), о которых было сказано чуть выше. И вот тут возникает вопрос! Если древние армяне могли себе позволить такие военные походы с Армянского нагорья на юго-юго-запад, то тем более не было смысла совершать подобные походы на запад, через Босфор в Европу? Армяне, добравшись до Европы, жили там долгое время (пару сотен лет), ассимилировали часть местного населения и впоследствии вернулись обратно, приведя с собой определенную часть европейцев в придачу к новым генам. Кроме того, длительное проживание на совершенно другой территории также может способствовать мутациям. Скорее всего, древние армяне совершили поход и в этом направлении, впоследствии «привезя» с собой на Армянское нагорье западноевропейский гаплотип R1b. P.S. В целом история армян достаточно прозрачна и бесспорна в своей основе до ~1000 г. до н.э. А сценарии «до этого» — бывают разные. 10 000 и более лет назад сформировалась наша нация. Есть аргументы в поддержку, хотя и не много. Правда, эти «осколки» порой собираются в очень четкие логические цепочки. Но, к сожалению, процесс глубокого познания осложняется многими факторами. 1. Армяне практически всю свою историю воевали, наши враги часто не гнушались попытками массового истребления, уничтожения культурного наследия. Многое было утрачено в результате войн, зависти соседей и жажды присвоения. Кстати, жажда присвоения существует и сегодня, например, в Грузии она обрела второе дыхание, об этом можно почитать здесь. Есть документы, фотографии… вот один из огромного количества примеров “грузинизации” армянских памятников из этого материала: 3. Сейчас нереально проводить раскопки на Армянском нагорье, туркам это ох как невыгодно… за последние 90 лет они уничтожили большую часть армянского культурного наследия в Западной Армении, а теперь еще и раскапывают новое… Пока там будут турки, ничего армянского там не найдут, а если и найдут что-то случайно, то все следы принадлежности к армянам будут уничтожены, или если это нереально, то сама находка будет уничтожена.
«Европейский палеолит» R1b сталкивается с непреодолимыми противоречиями. Последующие исследования
Барбары Арреди и коллег показали, что разнообразие субкладов этой гаплогруппы увеличивается по мере продвижения на восток, что скорее указывает на восточное происхождение этой гаплогруппы и связано с распространением сельского хозяйства из Западной Азии в Европу.
Этот вывод подтверждается дальнейшими исследованиями подгруппы R1b1a2, носителями которой могли быть фараоны Эхнатон и Тутанхамон. Предположительно она возникла на Армянском нагорье около 9500 лет назад — известно, что до того, как семиты поселились в Айасе и на Армянском нагорье, народы (урарты; хатты; хурриты) были широко распространены по всему Ближнему Востоку — и начали мигрировать в Европу около 7000 лет назад (см. Балканский неолит). Часть ее представителей мигрировала в Северную Африку (примером массовой миграции с Ближнего Востока в Египет является нашествие гиксосов, произошедшее за полтысячелетия до рождения Тутанхамона).

Происхождение и палеогенетика
R1b произошла от Y-хромосомной гаплогруппы R1, которая произошла от гаплогруппы R. Местом происхождения гаплогруппы R является Северная Азия. Период между 26 500 и 19 000 лет назад, в это время был последний ледниковый максимум. Древнейшей находкой Y-хромосомной гаплогруппы R* являются останки мальчика MA1 из стоянки Мальта в Иркутской области, возрастом 24 000 лет [13]. Популяция с Мальты произошла от популяции северо-восточных сибиряков (древние северные сибиряки, ANS), представленной жителями стоянки Янской с Y-хромосомной гаплогруппой P1, предковой для Y-хромосомных гаплогрупп Q и R [14].
Носители гаплогруппы R внесли большой вклад в происхождение современных жителей Европы и Южной Азии. В этих регионах наиболее распространена гаплогруппа R — R1b в Западной Европе, R1a в Восточной Европе, Центральной и Южной Азии и R2 в Южной Азии. Самые древние мутации R1b — M343, P25, L389 встречаются на огромной территории от Западной Европы до Индии [источник не указан 489 дней]. Это говорит о том, что мальтийский мальчик не оставил потомков в Сибири, а мигрант R-Y482 нашел свое убежище на Ближнем Востоке 28 200 лет назад [источник не указан 352 дня].
R1b1a2-V1636 (ISOGG2019-2020) был выявлен в энеолитическом образце ART038 (3356–3121 гг. до н.э.) из Арслантепе в Центральной Анатолии[15][16], в энеолитических образцах PG2001 и PG2004 (4233–4047 гг. до н.э.) из стоянки Прогресс-2 (Ставропольский край), у представителя ямной культуры SA6010 (2884–2679 гг. до н.э.) из могильника Шарахалсун 6 в Ставропольском крае[17], в образце Гьеррилд 5 (SGC/LN, 2283–2035 гг. до н.э.), который археологически относится к позднему неолиту, найденному в могильнике культуры одиночных могил Гьеррилд da [16].
Три основных субклада R1b1 — R1b1a, R1b1b, R1b1c — происходят с Ближнего Востока. Южная ветвь R1b1c — R1b1a2-PF6279/V88 встречается в Леванте и Африке. Северная ветвь R1b1a — R1b1a1a-P297/PF6398 возникла в Кавказском регионе, восточной Анатолии и, возможно, северной Месопотамии. Затем носители R1b1a1a-P297 переместились в Европу и Центральную Азию, разделившись на две ветви.
Взрывной рост численности потомков основателя субклада R1b1a1a2a1a-L52/P310/L11>PF6538>L151[en] произошел примерно 5,9–4,8 тыс. лет назад, что совпадает с распространением культуры колоколовидных кубков в Западной Европе и культуры боевых топоров в Восточной Европе.
Скотоводы неолитического периода[править | редактировать источник]
Ученые выдвинули теорию о том, что носители R1b (которые жили в неолите вместе с носителями J2) были первыми племенами, одомашнившими крупный рогатый скот[источник не указан 653 дня]. Генетический анализ ДНК современных коров и ее сравнение с ископаемыми образцами животных показали, что все современные коровы, которые произошли от дикого быка, одомашненного в неолитический и ранний сельскохозяйственный период, ведут свое происхождение от популяции из 80 животных, первых одомашненных коров. Самые ранние свидетельства одомашнивания коров датируются 10 500 годами и были обнаружены в гончарных культурах периода неолита, расположенных в горах южного побережья Тавра (хребта) на территории современной Турции. Древнейшие поселения со следами одомашнивания коров — Чайоню на юго-востоке Турции и на месте Джауд-эль-Мугхара на севере Ирака.
R1b1a2-V88 был выявлен в образце I5235 (9221-8548 гг. до н. э., Железные ворота, Падина, Сербия). R1b1a2b1-Y8451 был выявлен в образце R6 (7160 (7192–7127) лет назад, Апулия, Италия)
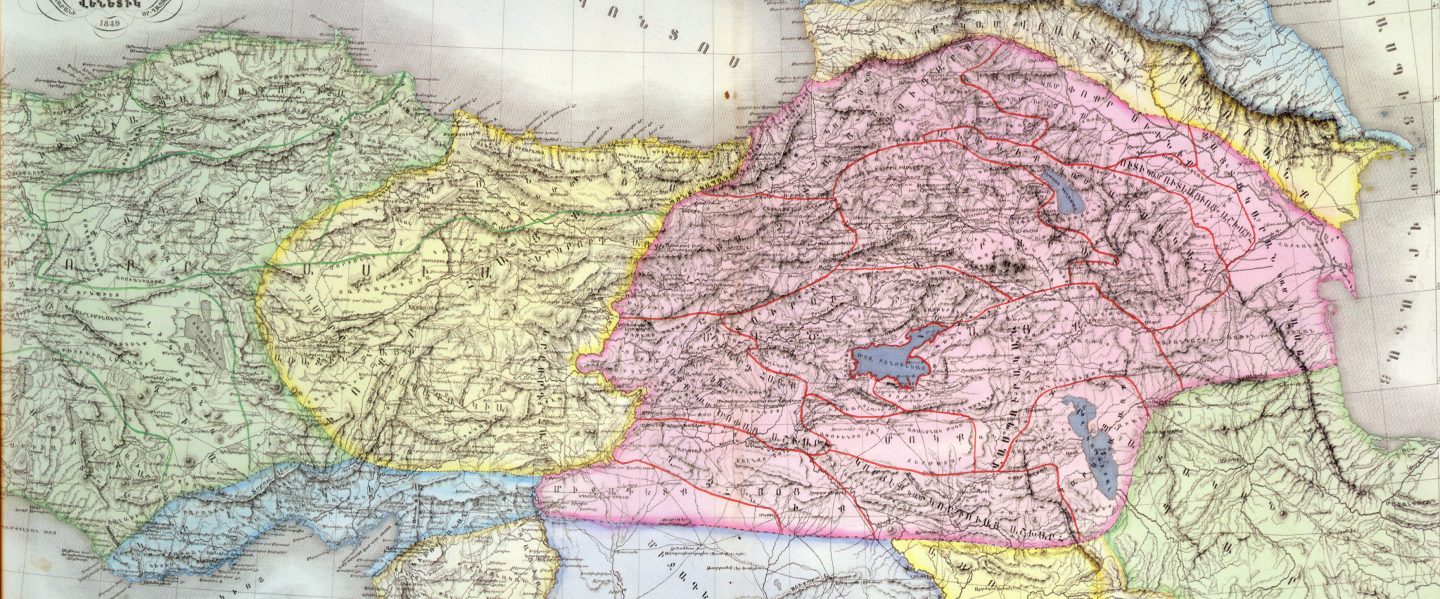
Армянское нагорье и Понтийско-Каспийская степь.

Современные лингвисты считают, что праиндоевропейцы зародились в Понтийско-Каспийских степях. Они простираются от устья Дуная до Уральских гор и до Северного Кавказа. Культуры неолита, энеолита и раннего бронзового века в Понтийско-Каспийской степи были названы единой курганной культурой, которая развивалась в период 6200 – 4200 лет назад, такое определение дала Мария Гимбутас в рамках своей курганной теории происхождения индоевропейцев. В современный период эта теория практически отвергается, в связи с тем, что курганы датируются не позднее 6000 лет назад и возникли на Южном Кавказе. Генетическое разнообразие R1b больше в Восточной Анатолии, этот факт говорит о том, что носители R1b эволюционировали там и южнее, а уже потом пришли на Армянское нагорье и попали в Понтийско-Каспийские степи.
R1b-P297 сформировался 15 600 лет назад (95% доверительный интервал 17 600 – 13 800 лет назад).
Лошади были одомашнены около 6600 лет назад в степях Прикаспия или на Дону или Нижней Волге. Этот прорыв был решающим для степных культур. Однако R1b не одомашнивали лошадей, они пришли в причерноморско-каспийские степи, когда эти животные уже были поставлены на службу людям. Вероятнее всего, лошадей одомашнили носители гаплогруппы R1a, вместе со старой ветвью R1b-P297, которая малочисленна по сравнению с R1a. R1b-P297 отнюдь не была индоевропейской, это были люди, которые еще жили в ледниковый период и не знали скотоводства. Эта ветвь поселилась в причерноморско-каспийских степях в позднем палеолите, что подтверждают находки в Самаре и Латвии. R1b-P297 и R1a по составу генома были практически чисто коренными жителями мезолитического периода, которые жили в Восточной Европе. В них была небольшая примесь сибирских охотников палеолита и мезолита, но не было примеси жителей Кавказа (CHG). В то же время примесь жителей Кавказа обнаружена в энеолитических культурах – афанасьевской, ямной и культурах шнуровой керамики. Гаплогруппа R1b1a1a2-M269 образовалась в позднем палеолите 13 300 лет назад (95% доверительный интервал 14 900 – 11 800 лет назад). Время до самого позднего общего предка (TMRCA) потомков R1b1a1a-M269 составляет 6400 лет назад, а непосредственно вышележащий узел R1b-P297 находится гораздо раньше 13 300 лет назад. Y-хромосомная гаплогруппа R1b-P297(xM269) была обнаружена у охотников-собирателей из Прибалтики, а также у охотников-собирателей из Самарской области России. «Длинная ветвь» R1b1a1a2-M269 значительно снижает уверенность в близости самых ранних носителей R1b1a1a2-M269 к этим восточноевропейским родственникам. Однако данные в равной степени согласуются со сценарием, в котором у основателя R1b1a1a2-M269 отсутствовал компонент восточного охотника-собирателя (EHG).
Точный маршрут миграции R1b1a1a2-M269 с Южного Кавказа в Понтийско-Каспийскую степь пока не ясен. Возможно, это произошло в днепровско-донецкий период, 7100–6300 лет назад. Эта культура стала первым неолитическим обществом в прикаспийской степи, у днепро-донецких носителей R1a и/или I2a1b уже были коровы и козы, пасущиеся в степи, и появились погребальные обряды, они также импортировали медь и медные изделия с территории Балкан и Карпат. Часть своих культурных традиций они переняли у охотников мезолитического периода. Знания о домашних животных пришли в эту культуру от земледельцев с Ближнего Востока, это были носители I, I2a2, R1a, R1b1a (L754) и одной R1b1a2-L388. Что важно, среди этих земледельцев не было носителей ветвей R1b1a1a2-M269 или низшей R1b-L23, которая тогда доминировала в ямной культуре. МтДНК днепро-донецкой культуры представлена исключительно женщинами европейского происхождения из мезолитического периода – U4a, U4b, U4d, U5a1, U5a2, U5b2, а также одной J2b1 и одной U2e1, все эти гаплогруппы мтДНК не являются индоевропейскими, а доиндоевропейскими. Вплоть до бронзового века индоевропейские мтДНК не встречаются в неолитических образцах, найденных на территории Украины. Таким образом, днепро-донецкая культура, которая уже имела как скотоводческие, так и сельскохозяйственные знания и была самой ранней такой культурой в степном Причерноморье, не принадлежит ни к индоевропейской культуре, ни к протоиндоевропейской культуре
Typical Indo-European mtDNA are — H2a1, H6, H8, H15, I1a1, J1b1a, W3, W4 and W5, these haplogroups of the female line will be found in the Yamnaya culture and the Unetice culture and then in other Indo-European cultures [24] [25]. The first Proto-Indo-European cultures were the Khvalynsk (7,200 – 6,500 years ago) and Sredny Stog (6,600 – 5,900 years ago). During this period, small burials of the kurgan type appear, with a characteristic position of the dead on their backs, with raised knees oriented to the northeast. In these cultures, the population actively entered into relationships with other tribes, this is noticeable by the various shapes of the skulls. 7000 years ago, the elite layer of these cultures began to actively develop using herds of cattle, horses and copper. Copper was a status metal, expensive and relatively difficult to mine and process. At the turn of the Khvalynsk and Sredny Stog cultures, a branch of R1b-L23 appeared from R1b1a1a2-M269, this happened approximately 6400 years ago (95% confidence interval 7100 – 5700 years ago) [33]. As a result, in the modern period, 99% of Indo-European R1b traces its roots to this branch – R1b-L23. Another clade R1b-PF7562 also originated from R1b1a1a2-M269, but it is small in number and is represented in the Balkans, Turkey and Armenia. The second migration through the Caucasus occurred about 5,700 years ago and coincides with the emergence of the Maykop culture, the world’s first Bronze Age culture. Most likely, the Maykop culture’s bearers came from Northern Mesopotamia, Syria and Eastern Anatolia. In the South Caucasus, the Leyla-Tepe culture (6,350 – 6,000 years ago) was discovered in Georgia and Azerbaijan. The ceramics of the Maykop and Leyla-Tepe cultures have similar features, as well as the dishes from the settlements of the Kura-Araxes culture. It is important that archaeological finds show a clear spread of burial mounds with bronze items from the Maykop culture in the Pontic steppe, it was in this steppe that the Yamnaya culture soon arose – about 5,500 years ago. The burial mound type became dominant and is one of the markers for determining the belonging of a culture to the Indo-European. Mounds as a method of burying nobles were then used by the Celts, Romans, Germanic tribes, and Scythians.
Период ямной культуры
Между 5500 и 4500 лет назад активно развивалась ямная культура бронзового века. Она считается бесспорно индоевропейской[источник не указан 408 дней] и важнейшей в основании индоевропейской культуры и общества. Носители R1b-M269 с Ближнего Востока в ямной культуре были относительно активны в отношениях с земледельцами и скотоводами R1a на протяжении более 2 тысяч лет, а также после ямной культуры в других сообществах. Эти контакты между R1a и R1b были не спорадическими, а плотными и многоуровневыми, как на уровне знати, так и на уровне простых людей и торговцев. Это привело к постепенному формированию уникальной общности населения ямной культуры, ямная культура имела относительно единый язык, который сегодня называют праиндоевропейским языком, который впоследствии стал основой практически всех современных европейских языков, включая все славянские языки, все англосаксонские языки и т. д. Лингвисты на современном этапе изучения сошлись во мнении, что именно регион понтийско-каспийских степей является родиной праиндоевропейского языка. Это подтверждается и сходством праиндоевропейского языка с кавказскими и хурритскими языками, происходящими с Ближнего Востока, а с другой стороны, с волго-уральскими языками. Только в Причерноморско-Каспийской степи пути носителей этих языков могли исторически пересечься настолько тесно, что в конечном итоге сначала возник бы праиндоевропейский язык — доиндоевропейский субстрат, затем индоевропейский язык [24][25][28].
Ямная культура наиболее ярко характеризуется тем, что ее народы использовали четырехколесные повозки для смены места жительства, перевозя на них продукты питания и предметы домашнего обихода. Такой транспорт обеспечивал возможность длительных походов по степям, что делало ямников одним из самых мобильных народов, одновременно занимавшихся земледелием и скотоводством. Этот набор навыков положил начало великим переселениям индоевропейцев. В более позднее время, в период Древнего Рима, свои миграции осуществляли также галлы и древние германцы [24][25][28].
Ямная культура, учитывая ее большие размеры, не была единой. На юге вдоль северного побережья Черного моря до Северного Кавказа располагалась полноценная степь, почти без леса, эта степь простиралась и дальше, до Каспийского моря, до Сибири и до Монголии и в англоязычной литературе называется Eurasian steppe.
1 — Западный ареал ямной культуры, между реками Дон и Днестр, а затем Дунай, был ареалом проживания преимущественно R1b, а носителей R1a там было не более 5-10% [источник не указан 408 дней].
Таким образом, западная часть ямной впоследствии мигрировала на Балканский полуостров и в Грецию. Затем в Центральный, и уже в сравнительно поздние исторические периоды — обратно, в виде волн миграций, состоящих из хеттов, фригийцев и армян, эти волны доходили до Анатолийского полуострова.
2 — Восточная часть ареала ямной культуры, от Волги до Уральских гор, была заселена R1a, а также меньшинством носителей R1b. Следы R1b сохранились в генетике некоторых башкирских родов, среди туркмен, уйгуров и хазарейцев.
Эта ветвь позже мигрировала в Среднюю Азию, и в Южную Азию — Иран, Пакистан, Индию
Европейская и ближневосточная ветвь
Когда индоевропейцы освоили бронзовое оружие и научились пользоваться конными повозками, их мобильность резко возросла. Они начали активно мигрировать в Европу, где в то время жили автохтонные, коренные культуры, носители C1a2, F и I (в том числе и те, кто несли прямые генетические связи с кроманьонцами). Также в Европе в начале индоевропейской экспансии жили неолитические земледельцы — земледельцы, мигрировавшие в раннем неолите и носительницы гаплогрупп G2a, H2, E1b1b и T1a. Индоевропейцы R1a и R1b, имея явное преимущество и в мобильности, и в вооружении, сумели за сравнительно короткое время полностью заменить мужские кланы в старых культурах Европы, скорее всего, они убили большую часть глав кланов или устранили их от возможности зачатия новых детей. При этом женская часть этих культур пострадала значительно меньше, что можно объяснить тем, что пришельцы R1a и R1b активно брали местных женщин в жены или наложницы.[25]
Если взять в качестве примера Южную Азию, то можно вывести следующее – после индоиранского (арийского) вторжения в Северную Индию 40% мужских линий стали R1a, но только 10% женских линий имеют индоевропейский корень, остальные женские линии остались коренными. Влияние индоевропейской экспансии на Европу было гораздо сильнее, в связи с меньшим развитием Европы 4000 лет назад, по сравнению с очень развитой Южной Азией в то время, где уже активно развивалась цивилизация Инда. В Европе 4000 лет назад еще не было бронзового оружия, земледелие было менее успешным. В Западной Европе земледелие появилось гораздо позже, чем на Балканах и в Центральной Европе.[25]
Греция, Балканы и Карпаты, с другой стороны, были наиболее продвинутыми регионами в Европе с точки зрения развития сельского хозяйства 4000 лет назад, и в результате их генетические линии были наименее затронуты индоевропейцами. Генетические гаплогруппы Старой Европы смогли выжить относительно нетронутыми только в регионах, где было неудобно путешествовать на лошадях – Альпы, Динарские Альпы, Апеннины и Сардиния.[25]
4700 – 4500 лет назад[править | редактировать источник]
Анализ ДНК культуры шнуровой керамики показывает, что уже 4700 лет назад ее носители в Польше имели как гаплогруппы R1a, так и R1b, но не субклад ямной культуры.
Движение R1b было направлено в сторону более развитой части Европы, населенной сельскохозяйственными культурами, и поэтому было более медленным. 4500 лет назад западная ветвь R1b набрала силу для новой мощной миграции в сторону современной Германии и Западной Европы. К этому времени носители R1b сильно смешались с обитателями бассейна Дуная, которые жили там до их прибытия, это были мезолитические охотники и неолитические земледельцы.[28]
R1b были явно патриархальным обществом. Они вытесняли местные мужские линии из культур, заменяя их R1b и активно брали женщин из этих культур в жены. Это привело к тому, что кожа протоиндоевропейцев стала темнее и по оттенку сравнялась с жителями современной Южной Европы, так как жители неолитических культур были выходцами со Среднего и Ближнего Востока и имели темные волосы и тон кожи.[25][28]
4500 – 3200 лет назад[edit | edit code]
R1b-L51 продвинулись в Центральную Европу (Венгрия, Австрия и Богемия) 4500 лет назад. Именно к этому времени, спустя 2000 лет после прихода неолитических культур – земледельческих, уровень их развития достиг высокого уровня, они начали добывать золото и медь и в конечном итоге стали гораздо более желанной добычей для завоевателей.
В настоящее время ставится под сомнение, была ли традиция колоколовидных кубков (4800 – 3900 лет назад) в Западной Европе полностью индоевропейской. Поскольку она является преемницей местных европейских культур, которые старше и относятся к мегалитическим культурам. В позднем неолите и раннем халколите колоколовидные кубки начали распространяться из Португалии на северо-восток, в сторону Германии. Степные культуры бронзового века, напротив, двигались из Германии в сторону Франции, Иберии и Великобритании и постепенно приносили R1b на территорию культуры колоколовидных кубков. Образцы индоевропейской керамики и конских фигурок, найденные в культуре колоколовидных кубков, скорее всего, являются не результатом индоевропейского компонента в этой культуре, а результатом торговли с носителями индоевропейского языка. Именно после этой торговли начался резкий рост миграции и завоеваний индоевропейских культур, и к 2200 г. до н. э. они достигли побережья Атлантики. Это может быть связано с тем, что культура колоколовидных кубков, когда она торговала, принесла индоевропейцам информацию о том, что в Западной Европе жили племена, которые еще не знали металла и находились на уровне каменного века – палеолита и поэтому были легкой добычей как для них самих, так и для территорий, на которых они жили [40][28]. ДНК-тесты носителей культуры колоколовидных кубков показали, что у них есть R1b-L51 (и более древние субклады – P312 и U152). Более ранние культуры до культуры колоколовидных кубков таких дополнений не имеют. Носители культуры колоколов, проживавшие на территории Испании и Португалии, не имеют примеси ямной культуры и не несли, за исключением R1b-L51, типичных гаплогрупп мегалитических культур, обитавших в Европе задолго до индоевропейцев, — G2a, I2a1, I2a2, а также R1b-V88, но эта ветвь не индоевропейская, а более древняя. [источник не указан 489 дней] Мегалитические культуры Великобритании относились исключительно к Y-гаплогруппе — I2 (в основном I2a2 и I2a1b-L161), но затем были полностью заменены R1b-L51 в период раннего бронзового века, что говорит о чрезвычайно интенсивной миграции ее носителей и, по сути, о полном завоевании и вытеснении местных жителей.
Гальштатская культура (между 3200 и 2750 годами назад) считается первой кельтской культурой в Европе — она быстро расширила свой ареал до Франции, Великобритании, Иберии, Северной Италии и долины реки Дунай. Гальштатская культура принесла с собой новый язык — кельтский, но бронзовая технология уже распространилась до нее.
Протоитало-кельто-германские племена достигли Германии около 4500 лет назад. Франция была заселена ими 4200 лет назад. Великобритания — 4100 лет назад. Ирландия — 4000 лет назад. Иберия — 3800 лет назад. Это была первая волна заселения, которая характеризуется линией — R1b-L21
Галльская и иберийская ветви
Первыми ветвями R1b, пришедшими во Францию, были R1b1a1a2a1a2c1-L21 и R1b1a1a2a1a2a-DF27[en]. 3700 лет назад эти линии достигли территории современной Испании. При изучении древних Y-хромосом была подтверждена активная экспансия R1b с изгнанием мужского коренного населения. В Иберии люди неолита и халколита были обладателями гаплогрупп I, I2a1 и G2a. А в бронзовом веке все мужчины уже были R1b (один M269 и два P312), но по данным мтДНК эти мужчины были сыновьями коренных иберийских линий — H1, U5b3, X2b. Развитие бронзовой культуры в Иберии было довольно медленным и бронзовый век, наконец, начался около 3300 лет назад в культуре полей урн, которая распространилась от Германии до Каталонии. После нее активно развивалась и расширяла свой ареал гальштатская культура (3200 – 2750 лет назад) – она заняла весь Пиренейский полуостров. К этому времени Иберия уже была связана с Западной Европой сложной торговой сетью. Начиная с 3300 лет назад вокруг Альп существовала область бронзовой металлургии, которая заложила основу для формирования классической кельтской культуры. Культуры в этой области создали преемственность – от культуры полей урн, которая развилась в гальштатскую культуру, начиная с 3200 лет назад, а затем гальштатская культура развилась в культуру Ла Тен – с 2450 лет назад. Это развитие представляет собой вторую волну экспансии носителей R1b в Европе, которая затронула Центральную Европу и двинулась в сторону Атлантического океана, северной Скандинавии и на восток в долину реки Дунай. Затем вторая волна достигла Греции, Украины и России.[43]
Кельтский железный век с 800 г. до н. э., возможно, возник в степях близ Северного Кавказа – кобанской культуры, которая существовала между 3100 и 2400 лет назад. У кельтов, населявших Альпы, была мутация S28 (другие названия – U152, PF6570)[44]
Гальштатская культура связана с мутацией S28. Эта культура позже развилась в виллановскую культуру (между 3100 и 2700 лет назад). Эта культура уже полностью индоевропейская, люди этой культуры помещали кремированные останки в урны в форме двойного конуса, захоронения делились на простые и на захоронения богатых людей и знатных людей. В знатных захоронениях содержались ювелирные изделия, бронзовые доспехи и конская сбруя. В этой культуре, среди прочего, появились типичные индоевропейские символы — свастика. За культурой Вилланова последовали этруски, демонстрируя прямую преемственность с этой культурой. У этрусков, проживавших в Центральной Италии с 800 г. до н. э. по 1 г. до н. э., с частотой 70% были выявлены Y-хромосомная гаплогруппа R1b (R1b-P312 и R1b-L2), с частотой 20% — Y-хромосомная гаплогруппа G2, а также Y-хромосомная гаплогруппа J2 (5%). Во времена Римской империи генетический вклад в этрусскую популяцию от лиц восточносредиземноморского происхождения увеличился примерно до 50%.
Germanic branch
Основная ветвь протогерманцев — R1b-S21 (она же U106 или M405).
Анатолийская ветвь
Фригийцы и протоармяне, скорее всего, пришли с Балканского полуострова. Они пришли в Малую Азию — Анатолию около 3200 лет назад в период «великих потрясений» в Восточном Средиземноморье. Фригийцы основали царство 3200 лет назад, оно просуществовало 500 лет и располагалось в центральной части Анатолии, в ее западном сегменте. Это царство поглотило большую часть распадающейся империи хеттов. Протоармяне пересекли Анатолию, достигли озера Ван и поселились на Армянском нагорье. В современный период 30% армян являются носителями R1b с субкладом L584 в Z2103. Большая часть R1b в современной Греции — это балканская разновидность Z2103.[источник не указан в течение 489 дней]. В Болгарии и Албании распространен субклад R1b-Z2106.
Другие миграции R1b
В современной популяции племени гайна R1b S28 (U152) встречается у 70% людей этой республики. R1b-S28 также является доминирующей ветвью на Апеннинском полуострове, в том числе в период Древнего Рима. Вероятнее всего, римские купцы во время походов по Великому Шелковому пути, который существовал уже 2200 лет назад, могли оставлять потомство от местных женщин, в том числе в регионе Средней Азии и Китая. Небольшой процент R1b есть среди населения христианских общин Ливана и, скорее всего, он остался от крестоносцев.
Этногеографическое распространение
Европа
Современная концентрация R1b наиболее высока в областях, связанных с кельтами: около 70% в южной Англии, до 90% и более в северной и западной Англии, Уэльсе, Шотландии, Ирландии, 70% в Испании и 60% во Франции [10]. По-видимому, она связана с докельтским субстратом, поскольку ее концентрация высока и среди некельтов: басков — 88,1% [источник не указан 876 дней] и испанцев — 70% [87]. Кроме того, известно, что, например, строителями Стоунхенджа в Англии были люди, которые жили на острове до прихода кельтов.
Концентрация этой гаплогруппы уменьшается среди соседних народов: 40% среди итальянцев, 40% среди немцев, 25,9% среди норвежцев [и других].
Еще реже она встречается среди народов Восточной Европы. Среди чехов и словаков – 22,6% [источник не указан 876 дней], поляков – 12,6%, латышей – 10,5%, венгров – 17,3%, эстонцев – 9%, литовцев – 5%, белорусов – 4,2%, русских – 7%, украинцев – 5,2%.
На Балканах – греки – 13,5% [95], словенцы – 21%, албанцы – 17,6%, болгары – 17%, хорваты – 11,7%, румыны – 13%, сербы – 9,6%, герцеговинцы – 3,6%.
Кавказ, Закавказье и Армянское нагорье
На Кавказе встречается у лезгин 21,5%, кумыков 25% и лакцев, аварцев (2,5%), андийцев, даргинцев (2%), кумыков, а также жителей Багулалы, осетин-дигорцев – по разным данным до 43%, у армян – 25-30%,
J2- Малая Армения Киликия и Армянское междуречье – Ряд современных генетиков считает, что J2 возник на Армянском нагорье. – Родина тех, у кого есть ген J2 – Малая Армения, Киликия и Армянское междуречье. J2 происходит от исходной oplogrupa R1b. Родина которой – Великая Армения!
R1b – Великая Армения – Ряд современных генетиков считает, что R1b возник на Армянском нагорье.
J2 – Малая Армения Киликия и Армянское междуречье – Ряд современных генетиков считают, что J2 возник на Армянском нагорье. – Родиной носителей гена J2 является Малая Армения, Киликия и Армянское междуречье. J2 происходит из исходной oplogrupa R1b. Родиной которой является Великая Армения!
Ряд современных генетиков считают, что J2 возник на Армянском нагорье.
Родиной носителей гена J2 является Малая Армения, Кликия и Армянское междуречье. J2 происходит из исходной oplogrupa R1b. Родиной которой является Великая Армения!
J2b был выявлен у представителя культуры Кура-Аракс
J2b2a-L283>CTS6190 был выявлен у этруска (образец R473, 700-600 гг. до н. э.).
J2b2a1a1a1b2~-Y86930 был выявлен у иллирийцев раннего железного века Хорватии (2765-2599 BP)
J2a1-L26>J-Y7010 был выявлен в образце I15543 (10 век) из Тимакума Минус (Равна, Кулине) в Сербии
J2b1-M205>J2b1a~-Y22075 был выявлен в образце BEL024 из славянского кургана № 96 (10-11 века) около деревни Студенка (Быховский район, Беларусь)
Распределение частоты J2 (включая все субклады): Малая Армения, Западная Армения, Восточная Армения, Ливан, Мальта, Италия, Греция, Иран, Грузия (особенно среди лазов и мингрелов), Македония, Сербия, Черногория, Беларусь, Россия (Русь, Осетия, Ингушетия и Чечня) и т.д.
Наконец, есть небольшая выборка иранских армян (Айоц Атрпатакан, Пайтакаран и Паркаик), которые, как полагают, были увезены в Исфахан (центральный Иран) войсками шаха Аббаса в 1604 году н. э.
Из этой выборки 25 человек показали 52 мутации на 0-5 шагах от ближневосточного гаплотипа, что дает время появления гаплотипа у предка этих людей как 2513,5 (2213,5) лет назад, а армянский гаплотип появился у них 5500 лет назад (17 человек с 25 мутациями на 0-5 шагах).
Естественно, при такой ограниченной статистике трудно рассчитывать на достоверные данные, но общий порядок появления двух основных гаплотипов у армян в целом совпадает – около шести тысяч лет назад
Ливанский тип армянской расы тип (по ближневосточному) гаплотип 14-15-23-10-11-12,
Армянский тип армянской расы – гаплотип 14-12-24-11-13-12. примерно три-четыре тысячи лет назад
Гаплотипы 12 колен как «основных» или «корневых» идентичны.
У армян один предок по мужской линии, который жил 6200 лет назад на Армянском нагорье (исторический ареал обитания армян по сей день).
Позже к ним присоединилась еще одна небольшая популяция с общим армянским предком. С тех пор генетическая картина в Нагорье не сильно изменилась.
Проект армянской ДНК на Family Tree DNA
Разные народы в разное время использовали разные названия для Армении и армян. Шумеры около 2800 г. до н. э. называли Армению Араттой, а аккадцы, пришедшие на смену шумерам во второй половине третьего тысячелетия до н. э., называли ее Армани или Арманум. Хетты, появившиеся во втором тысячелетии до н. э., использовали название Хайаса для обозначения Армении, а ассирийцы, возникшие во второй половине второго тысячелетия до н. э., называли ее Уруатри или Урарту (Арарат в Библии).
С исчезновением с исторической арены народов, называвших Армению и армян этими именами, исчезло и использование этих названий. Но Армения и армяне, которые всегда находили в себе силы и возможность обновляться на протяжении всего этого времени, продолжали и продолжают жить.
Аратта считается первым армянским государством, упомянутым в древних текстах. Это записи древнего шумерского эпоса о Гильгамеше. Считается, что этим текстам около 4800 лет, но это не значит, что государство Аратта не существовало до этого. Квентин Аткинсон и Рассел Грей показали, что армянский язык уже отделился от общего праиндоевропейского языка на Армянском нагорье, прародине индоевропейцев, около 8500 лет назад. Следует помнить, что древние армяне построили храмовый комплекс Портасар около 12 000 лет назад, что свидетельствует о высоком уровне социальной организации, безусловно, необходимом для осуществления столь грандиозной по меркам того времени работы.
Генетика и исследования происхождения армян
1) «В наших исследованиях предки армянского происхождения были прослежены в разных частях Армянского нагорья — Армении — Р.Армении, Р.Арцаха, Грузии, Азербайджана и Турции, что иллюстрирует тот факт, что историческая Армения была гораздо большей территорией, чем нынешняя Республика Армения».
Институт молекулярной и клеточной биологии Кафедра эволюционной биологии Урмас Русталу М.Н. RVA-Лиз Лугвали Проф. д-р Ричард Виллемс Тарту 2004
Original text in English
Происхождение армян и генетические исследования
1) «В нашем исследовании происхождение армян было прослежено до разных частей Армении, Грузии, Азербайджана и Турции, что иллюстрирует тот факт, что историческая Армения была гораздо большей территорией, чем нынешняя Республика Армения».
Тартуский университет Факультет биологии и географии Институт молекулярной и клеточной биологии Кафедра эволюционной биологии Урмас Роосталу М.н. Рва-Лийс Лоогвали Проф. д-р Ричард Виллемс Тарту 2004
Армяне принадлежат к 13 различным генетическим группам, возраст которых насчитывает десятки тысяч лет, и в то же время в ДНК армян за последние 4000 лет нет следов захватчиков, что делает их «однородными во всем их многообразии»
Генетика и исследования происхождения армян
1) «В наших исследованиях предки армянского происхождения были прослежены до разных частей Армянского нагорья — Армении — Р.Армении, Р.Арцаха, Грузии, Азербайджана и Турции, что иллюстрирует тот факт, что историческая Армения была гораздо большей территорией, чем нынешняя Республика Армения».
Институт молекулярной и клеточной биологии Кафедра эволюционной биологии Урмас Русталу М.Н. RVA-Лиз Лугвали Проф. д-р Ричард Виллемс Тарту 2004
Оригинальный текст на английском языке
Происхождение армян и генетические исследования
1) «В нашем исследовании происхождение армян было прослежено до разных частей Армении, Грузии, Азербайджана и Турции, что иллюстрирует тот факт, что историческая Армения была гораздо большей территорией, чем нынешняя Республика Армения».
Тартуский университет Факультет биологии и географии Институт молекулярной и клеточной биологии Кафедра эволюционной биологии Урмас Русталу Мн. наук Рва-Лийс Лоогвали Проф. д-р Ричард Виллемс Тарту 2004
Армяне принадлежат к 13 различным генетическим группам, возраст которых насчитывает десятки тысяч лет, и в то же время в ДНК армян за последние 4000 лет нет следов захватчиков, что делает их «однородными во всем их многообразии»
Анализ армянской ДНК.
1) Институт молекулярной и клеточной биологии Кафедра эволюционной биологии Урмас Русталу Мн. RVA-Liz Lugvali Проф. д-р Ричард Виллемс Тарту 2004 «В нашем исследовании предки армян были прослежены до разных частей Армении, Грузии, Азербайджана и Турции, что указывает на то, что историческая Армения была гораздо большей территорией, чем нынешняя Республика Армения».
2) Баной, Чалештори, Саньяти, Шариати, Хаушман, Маджидзаде, Солтани и Голиапур (2007) Изменение аллелей и генотипов DAT1 VNTR среди старых этнических групп в Месопотамии до региона Окс. «Армяне — это нация и этническая группа, происходящие с Кавказа и восточной Анатолии, где наблюдается большая концентрация этого сообщества, особенно в Армении».
3) (Мовсесян и Кочар 2000) «Сходство черепов современных армян и жителей Армении с 1600 по 700 гг. до н. э. указывает на непрерывность генетической связи с древним населением»
4) «Армяне — это отдельная этническая группа, которая произошла от неолитических племен Армянского нагорья» Литвинов С*, Кутуев А, Юнусбаев В, Р. Хусаинова, Валиев Р, Хуснутдинова Э.
5) 40% генов армян относятся к эпохе палеолита. Левон Епископосян
6) Армяне относятся к 13 различным генетическим группам, возраст которых исчисляется десятками тысяч лет, и при этом в ДНК армян за последние 4000 лет нет следов захватчиков, что делает их «однородными во всем их многообразии».
«В результате тестирования более 300 человек было установлено, что армянские ветви ДНК являются основой многих ветвей Европы».
Самоназвание Ари, слово «Ари». Ни один народ в мире не имеет такой прочной связи с этим понятием и словом, как АРМЯНЕ/АРМЕНЫ. Более того, ни в одном другом языке слово «Ари» не имеет ТАКОЙ ЧЕСТИ («Ари» = «храбрый», «Ари» = «мужественность», «КаджАри» = «мужественный», «Ар» = «мужчина», «Артакарг» = «супер» и т. д.), ни в одном другом языке мира нет столько названий с корнем АР (АРАРат — священная гора армян, Арагац, АРГейос, царство АРАрат; провинции Великого Айка — АРАРАТ, АРКАХ, округ Арк, АРТАЗ, АРАГАЦОТН; города АРМАВИРА, АРТАШАТ, АРАбкир, АРДАГАН, АРДВИН, АРЕШ, АРЧЕШ, АРРЗУМ, АРЦАП, реки АРАКС, АРАЦАН, крепость АРТАГЕРС и т. д.) и ни в одном другом языке мира нет столько личных имена с корнем «АР» (Армен, Арман, Артак, Арам, Аргам, Ара, Арташес, Арутюн, Артавазд и многие другие).
Первые арийские цари, чьи имена сохранились в Авесте, имеют явно армянские имена. Итак – Айомарт (дословный перевод с армянского – «армянский человек»), Айосинга (в переводе просто означает «Сын Айи», сын предыдущего), Аждаа (по-армянски означает «Великан»), Тиран-Тарон (/Траэтаона/, Тиран и Тарон – типично армянские имена), Манучар (опять же типично армянское имя), Назар (то же самое), Аршасп («Аршам», типично армянское имя).
И в этом вопросе огромное количество связей. Не говоря уже об огромном количестве свастик/солярных знаков на армянских храмах, церквях, доспехах и оружии воинов.
Кстати, забыл самое главное. АРев – солнце, а также два популярных символа – орел и лев – АРцив и АРуйц соответственно.
История происхождения R1b от ледникового периода до начала гальштатского периода (1200 г. до н. э.)
R1b — гаплогруппа Y-хромосомы, наиболее распространенная в Западной Европе. Она также встречается в Центральной Азии, Восточной Европе, Северной Африке и Западной Азии. После миграции европейцев в Америку и Австралию она составила там значительную долю. Она определяется однонуклеотидным полиморфизмом M343, открытым в 2004 году[4]. С 2001 по 2005 год R1b определялась наличием SNP P25. В более старых системах классификации это были Hg1 и Eu18.
Она наиболее распространена в Западной Европе, а также в некоторых частях России, таких как Дагестан. Гаплогруппа присутствует на более низких уровнях в Восточной Европе и в некоторых частях Северной Африки.
Она происходит от гаплогруппы R1, как и гаплогруппа R1a.
Гаплогруппа R1b возникла на Армянском нагорье из мутации гаплогруппы R1, произошедшей у человека, жившего примерно 22 800 лет назад (дата определена по фрагментам YFull). Последний общий предок носителей R1b жил 20,4 тыс. лет назад



Два важных субклада гаплогруппы R1a1a, по-видимому, широко разделяют европейскую и азиатскую части этой большой клады: R-Z283 (R1a1a1b1), по-видимому, охватывает большую часть R1a1a, обнаруженных в Европе (Pamjav 2012), в то время как R-Z93 (R1a1a1b2)
Эти два субклада происходят от гаплогруппы R1a1a (M17)/(M198), которая составляет подавляющее большинство всех R1a по всему ее географическому ареалу. R1a1a сама происходит от гаплогруппы R1a1 (SRY1532.2), которая возникла как единичная мутация одного мужчины, создателя R1a1, считающегося предком всех людей, несущих R1a1.
Гаплогруппа R
Считается, что гаплогруппа R (M207) возникла около 20 000–34 000 лет назад (Karafet 2008) где-то на Армянском нагорье, где ее предок гаплогруппа P-M45 чаще всего встречается в полиморфных частотах (Wells 2001). Древняя ДНК доказала, что ей не менее 23 000 лет.
Гаплогруппа R встречается на всех континентах, но довольно распространена в Европе, Южной Азии и Центральной Азии. Небольшие частоты встречаются в Малайзии, Индонезии, на Филиппинах и у коренных австралийцев (Kayser 2003). Она также встречается на Кавказе, на Ближнем Востоке, в некоторых частях Африки.
Два в настоящее время определенных субклада — R1 (M173) и R2 (M479). Гаплогруппа R1 имеет возможное время происхождения 26 000 лет назад, в то время как гаплогруппа R2 оценивается в 12 000 лет.
Происхождение R1 остается неясным. Гаплогруппа R является частью семейства гаплогруппы P (M45) и, следовательно, родственным кладом гаплогруппы Q (M242), которая распространена в Америке и Евразии.
На основании этих предковых линий, предполагаемое происхождение R1 к востоку от Западной Азии. Например, Kivisild 2003 полагает, что доказательства «предполагают, что южная и западная Азия могут быть источником этой гаплогруппы» и «Учитывая географическое распространение и разнообразие STR сестринских клад R1 и R2, последняя из которых ограничена Индией, Пакистаном, Ираном и югом Центральной Азии, возможно, что южная и западная Азия были источником дифференциации R1 и R1a».
Soares 2010 в своем обзоре литературы посчитал, что аргументы в пользу южноазиатского происхождения являются наиболее сильными, а центральноазиатское происхождение, аргументированное (Wells 2001), также заслуживает рассмотрения.
Гаплогруппа R2 определяется наличием маркера M479. Парагруппа для линии R2 обнаружена в Пакистане, Португалии, Испании, Италии, а также среди и осетин на Северном Кавказе.
Translation result
Гаплогруппа R1a (M420), вероятно, ответила от R1* примерно во время последнего ледникового максимума 19 000–26 000 лет назад. Мало что известно о месте ее происхождения. Некоторые считают, что она возникла на Балканах или в окрестностях Пакистана и северо-западной Индии из-за большего генетического разнообразия, обнаруженного в этих регионах. Гаплогруппа R1a1 (SRY1532.2) определяется SRY1532.2, также называемой SRY10831.2. Мутации SNP, которые, как считается, всегда происходят с SRY1532.2, включают SRY10831.2, M448, L122, M459 и M516. В этом семействе линий преобладает ветвь R-M17, которая положительна для M17 и M198. Парагруппа R-SRY1532.2* положительна для маркера SRY1532.2, но не имеет маркеров M17 или M198. Парагруппа R-SRY1532.2*, по-видимому, менее редка, чем R1*, но все еще относительно необычна, хотя она была протестирована в более чем одном опросе. Например, Underhill 2009 сообщает о 1/51 в Норвегии, 3/305 в Швеции, 1/57 греко-македонцев, 1/150 иранцев, 2/734 этнических армян и 1/141 кабардинцев. (Underhill 2009) В то время как Sahoo 2006 сообщил о R-SRY1532.2* для 1/15 образцов Химачал-Прадеш Раджпут (Sahoo 2006). Гаплогруппа R1a1a (M17)/(M198) составляет подавляющее большинство всех R1a по всему своему географическому ареалу. Она определяется мутациями SNP M17 или M198, которые всегда появлялись вместе у одних и тех же мужчин до сих пор. Два важных субклада гаплогруппы R1a1a, по-видимому, в целом разделяет европейскую и азиатскую части этой большой клады: R-Z283 (R1a1a1b1), по-видимому, охватывает большую часть R1a1a, обнаруженных в Европе (Pamjav 2012), в то время как R-Z93 (R1a1a1b2), по-видимому, охватывает большую часть R1a1a найдено в Азии. Современное распространение R-M17 отличается. Существуют две широко разнесенные области высокой частоты: одна в Южной Азии, вокруг Индии, а другая в Восточной Европе, вокруг Польши и Украины. Демографические причины этого являются предметом постоянного обсуждения и внимания среди популяционных генетиков и специалистов по генетической генеалогии, однако такие закономерности могут быть совокупным результатом (i) миграций и смешивания, (ii) естественного отбора и (iii) случайности. генетический дрейф.
Несмотря на заслуженную критику большинства археологов и антропологов, даже выдающиеся историки и археологи недавно попытались «поженить» доказательства из социальных наук с данными генетической антропологии. Хотя представление о том, что гены, язык и культура являются одновозрастными, весьма сомнительно, связь между R1a и «индоевропейцами» остается темой значительного научного интереса.
До 2012 года велись обширные научные дебаты относительно происхождения гаплогруппы R-M17. Это было результатом (i) отсутствия дальнейшего филогенетического разделения R-M17 на «дочерние» субклады и (ii) очевидно ошибочного убеждения, что мера «разнообразия STR» может однозначно квалифицировать, какая популяция имеет «самые старые» гаплогруппы R-M17.
Большая группа ученых обнаружила, что индийские или, в более общем смысле, южноазиатские популяции имеют самое высокое разнообразие STR. На основе этих исследований и с использованием эволюционно эффективной скорости мутации несколько из вышеупомянутых авторов пришли к выводу, что R-M17 присутствовал в южноазиатских популяциях со времен неолита, возникнув там.
Они также использовали эти доказательства для опровержения гипотезы о том, что R-M17 прибыл с индоевропейскими захватчиками с севера. Однако использование этой скорости мутации подверглось критике, поскольку ее не следует использовать с популяциями гаплогрупп, которые явно демонстрируют свидетельства расширения популяции, такими как R-M17.
Таким образом, использование этой скорости мутации может искусственно «раздуть» фактический возраст R-M17 в три раза. Действительно, авторы, использующие противоположное, «скорость мутации зародышевой линии» (которая является скоростью, эмпирически наблюдаемой в исследованиях отцов и сыновей), приходят к более поздним оценкам возраста. Фактически, Басби и др. недавно утверждали, что использование разнообразия STR при расчете «возраста» гаплогрупп весьма проблематично.
Другие исследования по-разному предполагали восточноевропейское, центральноазиатское и даже западноазиатское происхождение R – M17.
Десятилетний спор о том, какой евразийский регион обладал самыми разнообразными, а значит, и самыми древними значениями STR в пределах R-M17, был фактически положен конец с открытием подкладов R-M17. SNP предлагают более четкое и надежное разрешение, чем STR. Эти результаты на самом деле были известны в течение нескольких лет генеалогическим компаниям и генеалогам-энтузиастам, однако, наконец, были опубликованы две академические, рецензируемые статьи Памжавом и др. (2012, 2013). Они обнаружили, что все их протестированные индийские образцы R-M17 принадлежат к подкладу Z-93, который является производной, «дочерней» ветвью R-M17.
Напротив, восточноевропейские популяции принадлежат к разным дочерним ветвям R-M17, а именно Z-280 и M-458. Первый широко распространен в юго-восточной, центрально-восточной и восточной Европе, а также в Средней Азии. (Pamjav 2012) Действительно, Средняя Азия «является зоной перекрытия для R1a1-Z280 и R1a1-Z93», обнаруженных в монгольских и узбекских популяциях. С другой стороны, M-458 географически более ограничен центрально-восточной Европой.
Кроме того, это исследование показало, что недифференцированный, «родительский» M-198 существовал в европейских популяциях, но не был обнаружен в выборках индийских групп (состоящих из 256 малазийских индийцев, 301 цыгана, 203 дравидов из Индии).
Тем не менее, авторы пришли к выводу, что «Эта модель подразумевает, что ранняя зона дифференциации R1a1-M198, предположительно, произошла где-то в пределах евразийских степей или региона Ближнего Востока и Кавказа, поскольку они лежат между Южной Азией и Восточной Европой», откуда «южноазиатские» субклады Z-93 и «европейские» Z-283 дифференцировались и распространялись в противоположных направлениях.
Археологи признают комплекс взаимосвязанных и относительно мобильных культур, живущих в евразийской степи, часть которых простирается в Европу на запад до Украины. Эти культуры от позднего неолита и до железного века, с такими специфическими чертами, как курганные захоронения и одомашнивание лошадей, были связаны с распространением индоевропейских языков по всей Евразии.
Почти все образцы из могил бронзового и железного века в районе Красноярска на юге Сибири принадлежали к R-M17 и, по-видимому, представляли собой миграцию на восток из Европы.
В Центральной Европе останки человека периода шнуровой керамики в Эулау, из которых была извлечена Y-ДНК, по-видимому, относятся к R-M17(xM458) (который, по их мнению, наиболее похож на современный немецкий гаплотип R-M17*).
Происхождение
Гаплогруппа R1b (Y-ДНК) является доминирующей отцовской линией Западной Европы. В генетике человека гаплогруппа R1b является наиболее часто встречающейся гаплогруппой Y-хромосомы в Западной Европе и . R1b также присутствует с более низкими частотами по всей Восточной Европе, Западной Азии, Центральной Азии и в некоторых частях Северной Африки, Южной Азии и Сибири.
Из-за европейской эмиграции она также достигает высоких частот в Америке и Австралии. В то время как в Западной Европе доминирует R1b1a2 (R-M269). Они представляют собой две очень успешные «веточки» на гораздо большем «генеалогическом древе».
Ранние исследования были сосредоточены на Европе. В 2000 году Орнелла Семино и коллеги утверждали, что R1b была в Европе до конца ледникового периода и распространилась на север из иберийского убежища после последнего ледникового максимума. Оценки возраста R1b в Европе неуклонно снижались в более поздних исследованиях, по крайней мере, в отношении большинства R1b, причем более поздние исследования предполагают неолитический возраст или более ранний.
Только Морелли и др. недавно попытались защитить палеолитическое происхождение R1b1b2. Независимо от расчетов коалесценции STR, Чихи и др. указали, что время молекулярных расхождений не совпадает с расколами популяций; TMRCA гаплогруппы R1b (будь то в палеолите или неолите) датируется точкой ее происхождения где-то в Евразии, а не ее прибытием в Западную Европу.
Барбара Арреди и коллеги были первыми, кто указал, что распределение дисперсии STR R1b в Европе образует клин с востока на запад, что больше соответствует проникновению в Европу из Западной Азии с распространением земледелия. Статья 2009 года Кьярони и др. дополнили эту перспективу, используя R1b в качестве примера волнового распределения гаплогрупп, в данном случае с востока на запад.
Предположение о юго-восточном происхождении R1b было поддержано тремя подробными исследованиями, основанными на больших наборах данных, опубликованных в 2010 году. Они обнаружили, что самые ранние субклады R1b обнаружены в Западной Азии, а самые поздние — в Западной Европе.
Хотя оценки возраста в этих статьях все более поздние, чем последний ледниковый максимум, все они упоминают неолит, когда сельское хозяйство было завезено в Европу с Ближнего Востока, как возможный период-кандидат. Myres et al. (август 2010 г.) и Cruciani et al. (август 2010 г.) оба не определились с точной датировкой миграции или миграций, ответственных за это распределение, не исключая миграции в раннем мезолите или в конце Гальштата, но более вероятно в позднем неолите.
Они отметили, что для разрешения этих потоков генов могут потребоваться прямые доказательства из древней ДНК. Lee et al. (май 2012 г.) проанализировали древнюю ДНК человеческих останков из поздненеолитического памятника культуры колоколовидных кубков Кромсдорфа, Германия, идентифицировали двух мужчин, принадлежащих к гаплогруппе R1b Y. Анализ древней ДНК Y из останков популяций, происходящих из ранних неолитических поселений, таких как средиземноморский Кардиум и центрально- и североевропейские поселения LBK, выявил отсутствие мужчин, принадлежащих к гаплогруппе R1b.
Распространение
Река Днепр — одна из крупнейших рек Европы (четвертая по длине), берущая начало около Смоленска и протекающая через Россию, Беларусь и Украину к Черному морю. Общая длина составляет 2145 километров (1333 мили) с водосборным бассейном 504 000 квадратных километров (195 000 квадратных миль).
Река известна своими плотинами и гидроэлектростанциями. Днепр является важным судоходным водным путем для экономики Украины и соединен через Днепровско-Бугский канал с другими водными путями в Европе.
В древности река была известна грекам как Борисфен и была частью Янтарного пути. Археймар, столица готов, находился на Днепре, согласно саге о Гервараре.
Генетические исследования, основанные на современной мужской скандинавской ДНК, предполагают, что появление «культуры» Аренсбурга может быть связано с носителями гаплогруппы R1a1, которые, как предполагается, изначально распространились в Европу и принесли охотников на оленей «культуры» Аренсбурга из долины Днепра и Дона на Украине во время позднего ледникового максимума и достигли Скандинавии между 13 000 и 7600 лет назад.
Археологические свидетельства предполагают, что ядро древнейших популяций, колонизировавших Скандинавию 11 000–12 000 тысяч лет назад, пришло из современной Германии. Вероятно, они прошли через Ютландию и ныне затопленный Доггерланд, а затем направились на север через свободную ото льда прибрежную зону Норвегии.
Их каменные орудия позволяют предположить, что они принадлежали к аренсбургской культуре, которая процветала в Центральной и Восточной Европе (артефакты были найдены в Польше, Литве и России) в так называемый «поздний ледниковый период», то есть в конце LGM, когда повышение температуры и осадков вызвало отступление ледниковых щитов.
Анализ многих аренсбургских памятников и связанных с ними каменных орудий позволяет предположить, что эта культура зародилась в долине Днепра на Украине, одном из мест, где люди нашли убежище во время LGM.
Анализ полиморфизмов хромосомы Y в современных европейских популяциях показал, что Eu 19 (который также характеризуется другими маркерами хромосомы Y: 49a, fht11, SRY 1532G) распространился между 13 000 и 7600 лет назад из района долины Днепра и Дона, вероятно, когда группы, изначально искавшие убежища в этом районе во время LGM, получили возможность мигрировать из-за улучшения климатических условий в те регионы Европы, которые ранее были покрыты льдом.
Фактически, эта линия хромосомы Y встречается гораздо чаще в Восточной Европе с уменьшающимся градиентом на запад. Кроме того, она гораздо более разнообразна в популяциях Восточной Европы. Самая высокая степень диверсификации наблюдалась у украинцев.
Тогда возможно, что мужчины из Аренсбурга, а также большинство мужчин, происходящих из украинского убежища LGM, имели хромосомы Eu19 Y. Микросателлитные гаплотипы, связанные с M17 у норвежских индивидов, действительно представляют собой подмножество репертуара, наблюдаемого в Восточной Европе. В частности, наблюдалось преобладание гаплотипов 15.3/1 (21/19 повторов) и 16.5/1 (23/19 повторов) с их относительными производными.
Гаплотип Eu19 16.5/1 также очень распространен в Восточной Европе, в то время как гаплотип Eu 19 15.3/1 распространен в Норвегии, но очень редок в других местах. Этот своеобразный рисунок микросателлитов, связанных с EU19, может быть объяснен эффектом основателя, последующей изоляцией у норвежцев (и, возможно, у скандинавов) и в конечном итоге расширением локуса, как также наблюдалось в других местах.
Если кажется разумным предположить, что большинство мужчин Аренсбурга имели Y-хромосому Eu19, нельзя исключать, что они смешались с другими группами, прежде чем двинуться на север к норвежским берегам. В частности, позднеледниковая Центральная Европа характеризовалась экспансией северных балканских групп, где частота Y-хромосом M170 (EU 7), вероятно, была очень высокой.
Кроме того, основываясь на дифференциации гаплогруппы V в Скандинавии, также кажется, что группы, пришедшие из северного убежища Испании, вошли в Норвегию. Если это правда, то вполне вероятно, что Y-хромосомы M173 (EU18) также вошли в Норвегию во время позднеголедниковья.
Также считается, что R1a была доминирующей гаплогруппой среди северных и восточных носителей протоиндоевропейского языка, которые эволюционировали в индоиранскую, фракийскую, балтийскую и славянскую ветви. Протоиндоевропейцы произошли от культуры ямна (3300-2500 гг. до н. э.).
Эти популяции, вероятно, были носителями индоевропейского языка, которые, возможно, способствовали протобалтийско-славянско-германскому языковому единству в Прибалтике и северо-восточной Европе. В частности, культура шнуровой керамики из Центральной Европы (нынешняя Германия) и культура боевых топоров из Ютландии.
По всему Кавказу
Культура Телль-Халаф/Убейд связана как с Южным Левантом, так и с районом Закавказья и, возможно, вдохновила развитие как Кура-Акс, так и Майкопа. Район Кура-Акс и Майкопа становится ядром Циркумпонтийской металлургической провинции. У нас есть M269 L23xL51 на Кавказе и в Анатолии, а типы R1b L51 появляются у колоколовидных кубков, металлургов, в Западной Европе.
Палеолитическое происхождение R1b до сих пор не совсем ясно. Некоторые из древнейших форм R1b обнаружены вокруг Кавказа, в Иране и на юге Центральной Азии, обширном регионе, где могли бродить кочевые охотники-собиратели R1b во время ледникового периода. Ветвь R1 могла бы развиться в R1b, затем в R1b1 и R1b1a в северной части Ближнего Востока примерно во время последнего ледникового максимума (около 20 000 лет назад), в то время как R1a мигрировала на север в Сибирь.
R1b1a предположительно переместилась в северную Анатолию и через Кавказ во время неолита, где она разделилась на R1b1a1 (M73) и R1b1a2 (M269). Ближневосточные остатки эволюционировали в R1b1c (V88), который теперь встречается с низкой частотой среди ливанцев, друзов и евреев. Финикийцы (которые пришли из современного Ливана) распространили этот R1b1c в своих колониях, в частности на Сардинии и в Магрибе.
R1b1a2 (наиболее распространенная форма в Европе) и R1b1a1 тесно связаны с распространением индоевропейских языков, о чем свидетельствует их присутствие во всех регионах мира, где в древние времена говорили на индоевропейских языках, от Атлантического побережья Европы до Индийского субконтинента, включая почти всю Европу (кроме Финляндии), Анатолию, Армению, Европейскую Россию, южную Сибирь, многие районы вокруг Центральной Азии (особенно Синьцзян, Туркменистан, Таджикистан и Афганистан), не забывая при этом Иран, Пакистан, Индию и Непал. История R1b и R1a неразрывно связана друг с другом.
Современные лингвисты поместили прародину протоиндоевропейских языков в Понтийско-Каспийскую степь, особый географический и археологический регион, простирающийся от устья Дуная до Уральских гор на востоке и Северного Кавказа на юге. Неолитическая, энеолитическая и раннебронзовая культуры в степях Понтийского и Каспийского морей были названы Марией Гимбутас курганной культурой (7000-2200 гг. до н. э.) из-за устойчивой практики захоронения умерших под курганами («курганами») среди последовательных культур в этом регионе. Сейчас известно, что захоронения курганного типа датируются только 4-м тысячелетием до н. э. и почти наверняка возникли к югу от Кавказа.
Лошади были впервые одомашнены около 4600 г. до н. э. в степях Каспийского моря, возможно, где-то в районе Дона или нижней Волги, и вскоре стали определяющим элементом степной культуры. Тем не менее маловероятно, что R1b уже присутствовал в восточных степях в то время, поэтому одомашнивание лошади следует приписать коренным народам R1a.
Пока не совсем ясно, когда R1b перешел из восточной Анатолии в степи Понтийского и Каспийского морей. Это могло произойти во время или сразу после неолита, или и то, и другое. Поскольку генетическое разнообразие R1b было больше вокруг Кавказа, трудно отрицать, что R1b развился там до того, как войти в степной мир.
Возможно, что первая миграция R1b из Анатолии в 5-м или даже 6-м тысячелетии до н. э. принесла в степь овец, животных, шерсть которых будет играть важную роль в кельтских и германских (R1b ветви индоевропейцев) традициях одежды вплоть до наших дней. Другая миграция через Кавказ произошла незадолго до 3700 г. до н. э., когда майкопская культура, первое в мире общество бронзового века, появилось, по-видимому, из ниоткуда на северо-западе Кавказа.
Происхождение Майкопа до сих пор не определено, но археологи связывают его с современными энеолитическими культурами в Ассирии и западном Иране. Археология также показывает четкую диффузию бронзовой обработки и захоронений курганного типа из майкопской культуры в причерноморскую степь, где вскоре после этого (с 3500 г. до н. э.) развилась ямная культура (3500-2500 гг. до н. э.).
Ближневосточные люди R1b жили и в некоторой степени смешивались с местными собирателями и скотоводами R1a на протяжении более тысячелетия, возможно, даже двух или трех. Тесный культурный контакт и взаимодействие между людьми R1a и R1b по всей причерноморско-каспийской степи привели к созданию общего языка, нового лингва франка, который лингвисты назвали протоиндоевропейским (ПИЕ)
R1b1a2
R1b1a2 (название 2011 года) определяется наличием маркера SNP M269. R1b1a2* или M269(xL23) встречается с наибольшей частотой в центральных Балканах, в частности в Косово с 7,9%, Македонии с 5,1% и Сербии с 4,4%. Косово также примечательно тем, что имеет высокий процент потомков L23* или L23(xM412) в 11,4% в отличие от большинства других регионов со значительными процентами M269* и L23, за исключением Польши с 2,4% и 9,5%. Примечательно, что эта башкирская популяция также имеет высокий процент сестринской ветви M269 M73 в 23,4%. Пять человек из 110, протестированных в Араратской долине, Армения, принадлежали к R1b1a2 и 36 к L23*, при этом ни один из них не принадлежал к субкладам L23.
В европейском R1b доминирует R-M269. Он был обнаружен с низкими частотами по всей Центральной Евразии, но с относительно высокой частотой среди башкир Пермского края (84,0%). Этот маркер также присутствует в Китае и Индии с частотами менее одного процента. В таблице ниже более подробно перечислены частоты M269 в различных регионах Азии, Европы и Африки.
Частота составляет около 71% в Шотландии, 70% в Испании и 60% во Франции. В юго-восточной Англии частота этого клада составляет около 70%; в некоторых частях остальной северной и западной Англии, Испании, Португалии, Уэльса и Ирландии она достигает 90%; а в некоторых частях северо-западной Ирландии достигает 98%. Он также обнаружен в Северной Африке, где его частота превышает 10% в некоторых частях Алжира.
Как обсуждалось выше, в статьях, опубликованных около 2000 года, предполагалось, что эта клада существовала в Европе до последнего ледникового периода, но к 2010 году в центре внимания оказались более поздние периоды, такие как европейский неолит.
Диапазон новых оценок для R1b1b2, или по крайней мере его доминирующих частей в Европе, составляет от 4000 до максимум около 10 000 лет назад, и более детальное рассмотрение предполагает миграцию из Западной Азии через юго-восточную Европу. Западноевропейская R1b доминирует R-P310.
Также в этот период между 2000 и 2010 годами стало ясно, что особенно западноевропейская R1b доминирует определенными подкладами R-M269 (с некоторыми небольшими количествами других типов, обнаруженными в таких областях, как Сардиния).
В Европе R-M269 доминирует R-M412, также известный как R-L51, который, по данным Майреса и др. (2010) «практически отсутствует на Ближнем Востоке, Кавказе и в Западной Азии». Эта западноевропейская популяция далее делится на R-P312/S116 и R-U106/S21, которые, по-видимому, распространяются из западного и восточного бассейнов реки Рейн соответственно.
Myres et al. далее отмечают, что в отношении его ближайших родственников, в R-L23*, «поучительно», что они часто составляют более 10% популяции на Кавказе и некоторых юго-восточных европейских и циркумуральских популяциях. В Западной Европе он также присутствует, но в целом на гораздо более низких уровнях, за исключением «примера 27% в долине Верхней Роны в Швейцарии».
Кроме того, карта распределения подкладов, рисунок 1h под названием «L11(xU106,S116)», в Myres et al. показывает, что R-P310/L11* (или пока неопределенные субклады R-P310/L11) встречается только с частотой более 10% в Центральной Англии, а в прилегающих районах Англии и Уэльса частоты ниже.
Этот R-P310/L11* практически не встречается в остальной части Евразии и Северной Африки, за исключением прибрежных земель, окаймляющих западную и южную Балтику (достигая 10% в Восточной Дании и 6% в Северной Польше), а также в Восточной Швейцарии и окрестностях.
В 2009 году ДНК, извлеченная из бедренных костей 6 скелетов в раннесредневековом захоронении в Эргольдинге (Бавария, Германия), датируемом примерно 670 годом н. э., дала следующие результаты: 4 были обнаружены в гаплогруппе R1b с наиболее близкими совпадениями в современных популяциях Германии, Ирландии и США, а 2 были в гаплогруппе G2a.
Исследования населения, которые проверяют на M269, стали более распространенными в последние годы, в то время как в более ранних исследованиях мужчины в этой гаплогруппе были видны только в данных путем экстраполяции того, что вероятно. Ниже приводится краткое изложение большинства исследований, в которых конкретно изучался M269, показывающее его распространение в Европе, Северной Африке, на Ближнем Востоке и в Центральной Азии вплоть до Непала.
Армения
Армяне делятся на две основные группы в соответствии с их гаплотипами. Одна из них почти совпадает с атлантическим модальным гаплотипом (AMH):
Армянское нагорье является центром мировой цивилизации
Армянское нагорье имеет две большие гаплогруппы (R1b) и (J2), которые делятся на 13 подгоплогрупп.
Географическое распределение этого гаплотипа таково, что он разделяется армянами и двумя другими популяциями с Армянского нагорья. Более того, он отсутствует в большинстве других популяций с Кавказа, а также в других популяциях с более востока. С другой стороны, он чаще встречается в Европе, где, как мы знаем, гаплогруппа R1b также имеет тенденцию иметь более высокие частоты.
Было бы интересно тщательно изучить население современных Фракии, Анатолии и Армении и исследовать, может ли подгруппа хромосом R1b3, связанная армянским модальным гаплотипом, представлять собой сигнатуру обратной миграции в Азию балканских индоевропейских народов.
В сочетании с передовым бронзовым оружием и их морской культурой западная ветвь (R1b) индоевропейцев с берегов Черного моря являются прекрасными кандидатами на роль таинственных морских народов, которые совершали набеги на восточные берега Средиземноморья во втором тысячелетии до н. э.
Славяне и протоармяне — два других индоевропейских племени, происходящих с Армянского нагорья. Оба, по-видимому, мигрировали в Анатолию около 1200 г. до н. э. во время «великих потрясений» Восточного Средиземноморья.
Куро-араксская культура
Период между 4-м и 3-м тысячелетиями до н. э. был временем великих катаклизмов на Кавказе; его культурные достижения были обусловлены изменениями внутри его границ, а также взаимодействием с внешним миром. Самым значительным событием этой эпохи было появление большого количества народов месопотамской культурной идентичности, которые способствовали ускорению ритма его культурного развития, придавая «взрывной» характер его прогрессу.
В этот период армянское высокое пережило две мощные волны ближневосточной экспансии: первую во времена поздненеолитической культуры Сиони в 4-5-м тысячелетиях до н. э. и вторую в период культуры Цопи в поздненеолитическом веке, в конце 5-го и первой половине 4-го тысячелетия до н. э., которая известна как эпоха экспансии Урука.
Позднее, во второй половине IV и на протяжении III тысячелетия до н. э., в эпоху ранней бронзы, куро-араксская культура Кавказа распространилась по большей части Кавказа, Восточной Анатолии, северным районам Ирана, Ближнего Востока и даже Европы.
В этом контексте недавние археологические находки на Южном и Северо-Восточном Кавказе придали еще один, совершенно новый оттенок научным исследованиям древнего прошлого Кавказа. Они дали понять, что вторжение этих народов на Кавказ не было разовым событием, а продолжалось в течение довольно длительного периода.
Исходя из топографии археологических находок в Месопотамии, становится ясно, что большие массы переселенцев из этой местности не двигались прямо по маршруту в Закавказье, чтобы быстрее достичь цели. Фактически они селились в каждом регионе Кавказа, в горах и на равнинах, в местах, где они могли поддерживать привычный для них образ жизни.
Кажется очевидным, что с этого периода две культуры Кавказа, находившиеся на разных стадиях развития, могли мирно сосуществовать на основе взаимного участия в металлургическом производстве; именно этот тип общинного хозяйства дал толчок быстрому развитию местной культуры. Это хорошо иллюстрируется металлургическими изделиями куро-араксской культуры, которая значительно более продвинута по сравнению с доэнеолитической культурой.
В настоящее время ситуация кардинально изменилась. На основе целого ряда радиоуглеродных анализов доказано, что курганы древней ямной культуры относятся к значительно более позднему времени по сравнению с майкопскими археологическими памятниками.
Это позволяет ученым предполагать, что традиция сооружения такого типа курганов зародилась именно в майкопской культуре. Ее связи с левантийскими и месопотамскими древностями указывают на ее более раннее происхождение. В то же время, целый комплекс хронологических данных, полученных с помощью радиоуглеродного анализа, установил, что поселения и курганы Южного Кавказа, содержащие урукские артефакты, сосуществуют с майкопской культурой и, соответственно, древняя ямная культура и ее курганы относятся к более позднему периоду.
Поэтому сегодня мы не можем приписывать возникновение такого рода курганов в майкопской культуре, а также аналогичных одновременных памятников на Южном Кавказе влиянию культур степной зоны. Более того, не было никаких неблагоприятных условий, которые могли бы помешать появлению этого типа курганов на самом Кавказе
Ахундов (2007) недавно обнаружил докуро-араксские/позднеэнеолитические материалы из поселения Беюк Кесик и курганного некрополя Союк Булаг на северо-западе Азербайджана, а Махарадзе (2007) также раскопал докуро-араксский курган Кавтисхеви в центральной Грузии
Материалы, извлеченные из этих двух недавних раскопок, могут быть связаны с остатками из металлообрабатывающего позднего халколитического памятника Лейлатепе в Карабахской степи около Агдама (Нариманов и др. 2007) и с самым ранним уровнем многопериодного памятника Бериклдееби в Квемо Картли (Глонти и Дзавахишвили 1987). Они показывают наличие приподнятых могильных курганов или насыпей начала 4-го тысячелетия на Южном Кавказе.
Аналогичным образом, на основе своих исследовательских работ в восточной Анатолии к северу от Восточных гор Таурус, К. Марро (2007) сравнивает обожженные изделия, собранные в Ханаго на равнине Сюрмели и Астепе и Колпане в восточном районе озера Ван на северо-востоке Турции, с изделиями, найденными в вышеупомянутых местах, и связывает их с похожими изделиями (Amuq E/F), найденными к югу от гор Таурус в северной Месопотамии.
Культура Кура-Аракс или ранняя транскавказская культура была цивилизацией, существовавшей с 3400 г. до н. э. примерно до 2000 г. до н. э., что традиционно считается датой ее конца, но она могла исчезнуть уже в 2600 или 2700 г. до н. э.
Название культуры происходит от долин рек Кура и Аракс. Ее территория соответствует частям современной Армении,.
Об Армянох
Карта миграции людей Y-ДНК 2009 года, подготовленная Family Tree DNA, показывает, где возникли все гаплогруппы и как они распространились. Распределение гаплогрупп членов Армянского проекта ДНК (R1b, J2, J1, G, E, I1, I2, T, R2, Q, F, R1a, L, A) можно посмотреть на странице «Результаты». Связь между этими широкими гаплогруппами можно увидеть в этом ДЕРЕВЕ (ISOGG) и этом ДЕРЕВЕ (1000 геномов). Относительный возраст специфически европейских и азиатских гаплогрупп Y-ДНК можно увидеть в этом ДЕРЕВЕ, взятом из недавней статьи в Current Biology Педро Соареса и др. Эта армянская версия того же дерева дает вам представление о размере и разнообразии мужских гаплогрупп Y-ДНК в армянской популяции.
Карта миграции мтДНК человека 2006 года, подготовленная Family Tree DNA, показывает, откуда произошли все гаплогруппы и как они распространились. Это ДЕРЕВО дает представление об общей структуре ветвей. Распределение гаплогрупп членов Армянского проекта ДНК (H, U, T, I, W, HV, J, K, X, N, V, F) можно посмотреть на странице «Результаты». Относительный возраст европейских и западноазиатских гаплогрупп мтДНК можно увидеть в этом ДЕРЕВЕ, взятом из недавней статьи в Current Biology Педро Соареса и др. Эта армянская версия того же дерева дает вам представление о размере и разнообразии материнских гаплогрупп мтДНК в армянской популяции.
Модель африканского происхождения всех современных людей хорошо представлена в этом ДЕРЕВЕ, взятом из недавней статьи в Current Biology Майкла К. Кэмпбелла и Сары А. Тишкофф. Подробнее об этом. См. следующий раздел ниже на этой веб-странице.
Все участники проекта Армянской ДНК должны посетить веб-сайт Международного общества генетической генеалогии, чтобы просмотреть последнюю версию дерева гаплогрупп Y-ДНК ISOGG 2013. Нажав на букву своей гаплогруппы, они могут просмотреть самую последнюю информацию, касающуюся их ветви дерева (клады, подклады и т. д. с соответствующими мутациями SNP). SNP — это постоянные мутации ДНК, которые передаются всем мужчинам по прямой мужской линии. Они используются для определения ветвей внутри гаплогрупп.
Более новые, но гораздо более технические деревья Y-ДНК для некоторых гаплогрупп можно найти на веб-сайте DNA-Fingerprint, которым управляет Томас Крах, глава Центра исследований геномики FTDNA. Томас Крах также предоставил ссылку на самое современное (и техническое) дерево, доступное в форме DRAFT здесь. Ссылки из этого дерева на карту Y-хромосомы представляют интерес только для генетиков.
Family Tree DNA имеет другой способ представления деревьев для Y-ДНК в этом теперь уже датированном файле от 2009 года (большой файл).
Если вы столкнетесь со «старыми» статьями, использующими теперь уже устаревшую номенклатуру, вам может пригодиться это ДЕРЕВО от 2002 года (Ref. YCC NRY Tree 2002 v2002.01.18 от Консорциума Y-хромосомы в Университете Аризоны).
Argus Biosciences подготовила подробное филогенетическое дерево митохондриальной ДНК и КАРТУ, показывающую распределение гаплогрупп мтДНК по регионам в глобальном масштабе. Пожалуйста, увеличьте масштаб для лучшего просмотра.
Мнения, обсуждаемые в этом разделе, принадлежат Диенекесу Понтикосу в соответствии с его анализом недавней статьи Майкла ГБ Блума и Маттиаса Якобссона: «Глубокие расхождения генетических деревьев человека и модели происхождения человека». Как видно из этого рисунка, существует четыре или пять сценариев, объясняющих нынешнее распределение человеческих популяций.
R1b1a2a представляет собой самую большую гаплогруппу для армян в целом и участников проекта в частности. По оценкам, ей 8000 лет. По словам Винса Визакеро, который руководит проектом гаплогруппы R-ht35: «Из предыдущего анализа следует, что R1b1a2a довольно быстро двигалась на север и запад в Европу. И данные, которые мы видим в нашем проекте, согласуются с этим: самые старые формы R1b1a2a встречаются с высокой частотой на «родине — Армянском нагорье» Юго-Западной Азии и в местах с наибольшим контактом с этим регионом. Чем ближе мы приближаемся к Северо-Западной Европе, тем больше мы наблюдаем самые молодые, производные формы R1b1a2a». Текущее распределение этой гаплогруппы показывает сильную концентрацию в Западной Европе (от северной части Пиренейского полуострова до Ирландии и Англии через Францию и Бельгию), как можно увидеть на этой КАРТЕ. Карта подтверждает выводы Винса, поскольку она показывает 15% концентрацию R1b1a2a в северной полосе Анатолии – с пиком 25% в середине полосы. Исследования, на основе которых были составлены карты, охватывали широкий регион, включая турок, курдов, грузин и азербайджанцев. Если вы выберете только армян, вы получите концентрацию 30% R1b1a2. Если вы выберете только армян из Карабаха и Сюника, вы получите концентрацию более 40%.
Распределение “самых молодых и производных форм” R1b1a2, обнаруженных в основном в Западной Европе – включая всех армян на данный момент – можно посмотреть здесь (цифры представляют собой проценты). Обе являются единственными известными ветвями R1b1a2a1a1 (ht-15): P312 = R1b1a2a1a1a и U106 = R1b1a2a1a1b. Если в будущем в одной из этих двух групп будут обнаружены армяне, это, скорее всего, будет означать, что они произошли от крестоносцев. Согласно Википедии: Первоначально считалось, что R1b возникла в Западной Европе, где (рассматривая в целом, включая субклады) она достигает своих самых высоких частот. Однако дисперсия R1b увеличивается по мере продвижения на восток, что приводит к мнению, что R1b возникла дальше на восток и (M269) распространилась в Европу в неолите, а не палеолите. Многие генетики теперь считают, что R1b возникла в Армянском нагорье или Западной Азии. Все вышесказанное достаточно ясно проиллюстрировано на этом слайде PowerPoint.
Эта длинная статья о R1b1 довольно точна и актуальна. Рекомендуется к прочтению для членов этой гаплогруппы. Однако предупреждение относительно статей о других гаплогруппах: в них есть многочисленные неточности и упущения.
Винс Визакеро подготовил следующую КАРТУ распределения R1b1a2* (желтый – L265+), R1b1a2a* (зеленый – L23+), R1b1a2a1a* (оранжевый – L51+) и R1b1a2a1a1* (красный – L11+). Подавляющее большинство членов R1b1a2 Армянского проекта ДНК принадлежат к зеленой ветви R1b1a2a* L23+. Несколько являются частью желтой ветви R1b1a2* L265+. Ни один из них не является оранжевым R1b1a2a11* L51+. Один – красный R1b1a2a1a1* L11+. Что еще более важно, армяне не обнаружены в крупных европейских ветвях ниже красной L11+, а именно U106 и P312.
Недавнее исследование, опубликованное в январе 2010 года, похоже, подтверждает все вышесказанное. По словам авторов (Балареске и др.): «Гаплогруппа R1b1a2 является самой распространенной европейской Y-хромосомной линией, частота которой увеличивается с востока на запад, и ее носителями являются 110 миллионов европейских мужчин. Предыдущие исследования предполагали палеолитическое происхождение, но здесь мы показываем, что географическое распределение ее микросателлитного разнообразия лучше всего объясняется распространением из одного источника на Ближнем Востоке через Анатолию во время неолита… R1b1a2 была перенесена как быстро расширяющаяся линия с Ближнего Востока через Армянское нагорье на западную окраину Европы во время неолита. Наша интерпретация истории hg R1b1a2 теперь делает Европу ярким примером того, как расширение Y-хромосомной линии имеет тенденцию сопровождать технологические и культурные изменения».
Другое исследование, опубликованное в августе 2010 года (Myres et al.), подтверждает эту точку зрения: «Филогенетические связи многочисленных ветвей в пределах основной гаплогруппы Y-хромосомы R-M207 подтверждают западноазиатское происхождение гаплогруппы R1b, ее первоначальная дифференциация там сопровождалась быстрым распространением одного из ее подкладов, несущих мутацию M269, в Европу».
Обновление от января 2011 года: теперь, похоже, есть некоторая структура для крупнейшей ветви гаплогруппы в Армянском ДНК-проекте. Армяне и другие, проходящие тестирование в 23andMe, похоже, принадлежат либо к одному из двух новых L277 или L405. Масштабное тестирование начнется, как только эти мутации SNP станут доступны для тестирования в Family Tree DNA. Ссылка. это ясное новое ДЕРЕВО, подготовленное Винсом Визакеро.
Если все вышесказанное верно, это может объяснить происхождение басков.
Для последних исследований менее дифференцированной, но не обязательно более древней ветви гаплогруппы R1b1* посетите страницу новостей проекта ДНК R1b1* Винсента Визакеро. Визакеро вычислил длину ветвей для R1 и получил следующие оценки: R1 = 18 500 лет назад; R1b1 = 15 300 лет назад; и R1b1a = 12 500 лет назад. Вот сводное ДЕРЕВО для R1b1 и ее основных ветвей. Визакеро отмечает, что R1b1* существует по всей Евразии. Она довольно редка в Европе, и невозможно обойти тот факт, что R1b1 по сути является евразийской гаплогруппой с сильной филогенетической структурой. Согласно этой КАРТЕ, пик частоты R1b1* приходится на Юго-Западную Азию — там она, по-видимому, также имеет большее разнообразие.
По словам Винса: «В R1b1 есть две основные ветви (R1b, вероятно, фантом). Одна ветвь — это R-P297 [R1b1b], а другая — то, что мы называем R1b1 (или R1b1(xP297)). Так что если вы ищете точку происхождения для R1b1, то вы ищете пересечение этих двух множеств. TMRCA для этих двух линий составляет примерно 16 000 лет назад. R-P297 — это просто, поскольку есть только два клада, которые легко идентифицируются по SNP. R1b1a1 и R1b1a2 имеют евразийское распространение, а точка происхождения для обоих находится на Ближнем Востоке (назовите его Анатолией+Левант, если хотите). TMRCA R-P297 составляет примерно 12 000 лет назад. Хотя частота R-M73 [R1b1a1] самая высокая в Центральной Азии, это не то место, где он наиболее разнообразен. И есть три отдельных клада R-M73. R1b1* сама делится на две основные линии, разделение между которыми произошло примерно 15 000 лет назад. Ученые никогда не уделяют достаточного внимания R1b1, но мы видим, что пик частоты R1b1 явно находится на Ближнем Востоке.”
Первый кластер R1b1* (назовем его A) состоит исключительно из европейцев и армянина. Второй кластер R1b1* имеет две подгруппы (назовем их B1 и B2). B1 состоит из армянина, иракского сирийца и иранского араба. “B2 состоит из большой группы людей (включая европейцев, арабов, ашкенази, африканцев и т. д.). Именно эта группа B2, по-видимому, была исходной популяцией для миграции обратно в Африку в Камерун.” Кластер B1 отделился от кластера B2 около 10 000 лет назад. Новая мутация SNP под названием V88 отделяет A и B1 от B2. Винс Визакеро считает, что «R1b1 впервые появился в северной части юго-западной Азии (например, Ирак/Иран), тогда как V88 впервые возник в популяции R1b1, живущей немного южнее (например, Левант или Аравия)».
Члены гаплогрупп G2a и G1 могут узнать о различных кладах G на: категории гаплогруппы G, образцы, диаграммы и т. д. Они также могут узнать о временных связях и миграционных моделях гаплогруппы G. Вы можете копнуть глубже, изучив всестороннюю категоризацию образцов G вместе с некоторыми диаграммами. Эти сайты были созданы администратором проекта G DNA, Рэем Бэнксом, который поддерживает в актуальном состоянии страницы Википедии для: G, G1, G2a1, G2a3a и G2a3b1. Мутация гаплогруппы G возникла около 21 000–14 000 лет назад. Независимо от даты или конкретного места происхождения, часть семейства G пустила корни преимущественно в районе к югу и востоку от Кавказских гор. Сегодня Кавказ — это в основном страны Грузии, Армении, Арцаха и юго-западной России. Регион происхождения основных групп G сегодня, скорее всего, находился где-то в Армянском нагорье, регионе Кавказских гор, Иране или восточном Средиземноморье — во всех местах, где G встречается как в значительных количествах, так и с разнообразием, ожидаемым при длительном присутствии.
По словам Рэя Бэнкса, администратора проекта G в FTDNA: “G2a3a встречается в значительных количествах в Армянском нагорье, Греции и странах Восточного Средиземноморья. Люди G2a3a, по-видимому, распространяются на запад, в основном вдоль Средиземноморья из этих регионов. Очень предварительные расчеты показывают, что мутация M406, которая характеризует G2a3a, возникла около 2100 г. до н. э. в качестве очень общей оценки. Подробные образцы, доступные из внутренней Европы, сравнивались с подробными образцами из более восточных мест, а именно (1) Турции (2) Ливана-Иордании и (3) Армении. Эти сравнения показывают, что большинство европейцев имеют армян в качестве своих ближайших родственников, причем разделение с ними началось, как правило, около 1300 г. до н. э. и продолжалось в период Темных веков после Римской империи. Те, у кого было самое старое разделение (обычно около 1300 г. до н. э. – 800 г. до н. э.), показывают разделение со всей группой на востоке – в равной степени – а не с определенным регионом. Этого следовало ожидать, поскольку возраст мутации, вероятно, не простирается гораздо дальше назад». Некоторые исследователи полагают, что ветвь G2a3a1, возможно, возникла несколько тысяч лет назад в хурритском/урартском населении Армянского нагорья.
Истоки G1, по-видимому, находятся в Иране. Однако, поскольку было тщательно протестировано так мало G1, мы пока не можем сделать никаких однозначных выводов. Профессор Левон Епископосян обнаружил высокую концентрацию гаплотипов G1 у армяноязычного населения амшенов Абхазии (первоначально из Орду и, по предкам, с гор над Трабзоном/Трапезундом и Ризе).
Гаплогруппа G, по-видимому, распространилась из юго-западной Азии или Ближнего Востока в Европу к западу от Черного моря и в западное Средиземноморье и Центральную Азию в основном около 1700–3000 лет назад. Три страны — Грузия, Армения и Азербайджан — предоставляют образцы STR, соответствующие всем типам гаплогруппы G, которые сегодня встречаются в Европе к западу от Черного моря, за исключением G1 и G2c. Из-за богатого разнообразия этих типов G эта область мира является ведущим кандидатом на отправную точку для большинства миграций G, особенно в Западную и Центральную Европу.
Эта КАРТА показывает текущее географическое распределение гаплогрупп J1 и J2. Новая КАРТА J2a (M410+) была опубликована в декабре 2010 года. Армянские члены этих гаплогрупп в настоящее время принадлежат к следующим ветвям: J1, J1c3d, J1c3d1, J2a, J2a3, J2a4, J2a4a, J2a4b, J2a4b1, J2a4d, J2a4h2, J2a4h2a, J2a4h2f, J2a4h2g, J2b* и J2b1. Вот последние данные по черновой структуре ветвей J1 и J2. Это последнее дерево для J1.
Члены кладов гаплогрупп J1 и J2a4* могут просмотреть последние диаграммы Fluxus, созданные администратором проекта J DNA Бонни Шрак на этой странице проекта Y-Haplogroup J DNA. Они показывают, как различные ветви и кластеры внутри каждой группы соотносятся друг с другом.
Доктор Рой Кинг из Стэнфордского университета считает, что «J1* с DYS388 = 13 или 14, L136-, P58-субклад» является одним из маркеров кавказоязычных хурритов и урартов. Джеймс Ханичак подготовил КАРТУ текущего распределения этого субклада. Многие, если не большинство восточноевропейских случаев J1* с DYS388 = 13 или 14, показанных на этой карте, вполне могли быть предками армян.
Члены ветви J2a4h (L25) принадлежат к двум различным кластерам. Более старый из них характеризуется значением 10 в DYS445, более молодой — значением 6. По словам Дэвида Дугаса: «В настоящее время мы думаем, что 445=10 является предковым состоянием для этого субклада, и его источником может быть Ирак или регион, где встречаются Армянское нагорье, Иран и Ирак. 445=6, нижестоящий кластер, частично определяемый L70, возник в левантийском или прибрежном анатолийском регионе из гаплотипа 445=10». Вот что говорит Аль Абурто о младшей ветви: «Не совсем ясно, где и когда первый человек, уже имеющий L25, имел первую делецию DYS445=6 из состояния DYS445=10 (DYS445=10 наиболее распространен у остальных людей L25). Вероятно, это произошло в Южноармянском нагорье, Северной Сирии или Ливане — где-то в этом районе. Временные рамки также не ясны, но из программы, которую я использую (Generations5), кластер L25(xL70) с DYS445=6, похоже, имеет возраст около 3600+/-400 лет. Эта КАРТА показывает, где были обнаружены гаплотипы L25*(6) до сих пор. Данные с Ближнего Востока (которые прояснят происхождение) постепенно накапливаются». Эти данные недавно позволили администраторам проекта подготовить эту КАРТУ дисперсии.
Члены одной из старейших гаплогрупп, E1b1b1, могут просмотреть маршрут, по которому она распространилась из Африки, а также предполагаемый относительный возраст различных субкладов. Вот последнее ДЕРЕВО ветви E1b1b1… а вот ДИАГРАММА для альтернативного взгляда.
Гаплогруппа E, по-видимому, возникла в Северо-Восточной Африке, основываясь на концентрации и разнообразии субкладов E в этой области сегодня. Но тот факт, что гаплогруппа E тесно связана с гаплогруппой D, которая не встречается в Африке, оставляет открытой возможность того, что E впервые возникла на Ближнем или Среднем Востоке и впоследствии была перенесена в Африку обратной миграцией.
Армянские члены проекта относятся к V12, V13, V22 и M84/M34. Связь между этими тремя большими субкладами E1b1b1 = M35 можно увидеть на этой диаграмме M215+. Считается, что E1b1b1 = M35 возник 25 000 лет назад.
Он известен как E1b1b1c1, возник в Анатолии. Оттуда он распространился на запад в Европу и на юг на Ближний Восток. Разнообразие гаплотипов наиболее высоко в Плодородном полумесяце, уменьшаясь к Аравийскому полуострову.
E1b1b1a2, берет свое начало на Ближнем Востоке или в Армянском нагорье.
MD: Посмотрите на эту карту, которая, как я считаю, несколько неполна. Эту статью Стивена Берда о V13 обязательно нужно прочитать. Некоторые линии Y-ДНК могли быть введены в армянское население таким же образом римскими вспомогательными легионерами, прибывшими на Южные Балканы. появиться… но структура остается неясной, как и местоположение V13. Мнение о V13 заключается в том, что она возникла где-то на севере Леванта и на последней границе Армянского нагорья.
Гаплогруппы I1 и I2 являются потомками супрагаплогруппы F, которая, как полагают, представляет собой вторую и более позднюю фазу миграции человека из Африки 50 000 лет назад. Как видно на этой карте, гаплогруппа I и ее сестра считаются кладами J1 и J2. достигли Европы через Ближний Восток, используя Левантийский коридор между Тигром, Евфратом и Евфратом. является частью древнего плодородного полумесяца, образованного речными системами. Реки Нил. Следует отметить, что гаплогруппа J присутствует и очень характерна для Ближнего Востока, в то время как гаплогруппа I здесь почти не существует и, похоже, характерна исключительно для Европы, как видно на этой карте (M253 = I1; M26 = I2a1; M423 = I2a2 ; M223 и M284= I2b1).
Гаплогруппы I1 и I2 считаются потомками супрагаплогруппы F. Как видно на этой карте, гаплогруппа I и ее родственные клады J1 и J2, как полагают, достигли Европы через Ближний Восток, используя Левантийский коридор, который является частью древнего плодородного коридора. полумесяц, образованный речными системами Тигр, Евфрат и Евфрат. Реки Нил. Следует отметить, что гаплогруппа J присутствует и очень характерна для Ближнего Востока, в то время как гаплогруппа I здесь почти не существует и, похоже, характерна исключительно для Европы, как видно на этой карте (M253 = I1; M26 = I2a1; M423 = I2a2 ; M223 и M284= I2b1).
По словам Кена Нордтведта, специалиста по гаплогруппе I: «За 12 000 лет до нашей эры [до настоящего времени] Европа и близлежащие регионы пережили длительный холодный период продолжительностью более тысячи лет — Поздний дриас. Вероятно, это был последний серьезный демографический спад, который пережили наши предки по всей Европе. Хотя гаплогруппа y I (y-Hg I) к тому времени была зрелой гаплогруппой, которой, возможно, уже 10 000 лет, я пришел к выводу, собрав и изучив от пяти до десяти тысяч гаплотипов y-Hg I на сегодняшний день, что только девять мужчин вышли из Позднего дриаса с выжившими потомками по мужской линии на сегодняшний день. Эти Девять Поздних Дриасов теперь имеют десятки миллионов потомков мужского пола в Европе и других местах земного шара, где европейцы поселились в последние столетия: каждый из этих девяти мужчин был не одинок; мужское население охотников-собирателей из ближайших родственников, расширенной семьи: семьи, клана, племени и т. д. Некоторые из этих соседних мужчин несли y гаплотипы, очень близкие к одному из девяти и произошедшие от общих предков, не намного более отдаленных во времени. Эти клады гаплотипов, окружающие каждую из девяти, можно было бы насчитать десятками, сотнями, может быть, даже тысячами современников. Но из-за очень высокой вероятности вымирания для этих мужских линий, превышающей 99 процентов, эти девять счастливчиков появились как единственные представители своих кладов, имеющие сохранившиеся линии сегодня. Многие y-клады, без сомнения, полностью вымерли в ту эпоху». Вы можете прочитать весь пост Кена здесь.
Остается выяснить, как / куда вписываются армяне, являющиеся членами этой гаплогруппы. Изначально я думал, что они будут либо остатками изначального анатолийского населения, либо потомками европейцев, которые мигрировали обратно на Армянское плато. Оба случая возможны. Есть соблазнительные намеки на то, что I2* возникла в этом регионе, поскольку до сих пор она была обнаружена только в Армении, Армянском нагорье и на Кавказе. По словам Кена, «Армения становится горячей точкой за пределами Европы для I2. Туман, возможно, рассеивается относительно того, где именно IJ породил I и J” и “I2 … является единственным кладом или подгаплогруппой Hg I с вероятным коренным населением за пределами Европы”.
Также по словам Кена (отредактировано): “F-man, безусловно, является предковым узлом для узла IJK, а затем узла IJ и позже узла I. Мы никогда не находили ни одного гаплотипа IJK, IJ или I. И между узлом IJK и самыми ранними кладами “I”, которые мы видим, проходит очень много времени — вероятно, I2 и I2a1 M26+. Мы говорим о нескольких десятках тысяч лет, что, безусловно, указывает на Армянское нагорье. «Ребята с Армянского нагорья двинулись на запад, к северу или югу от Черного моря. Я сейчас делаю ставку на южный маршрут, но без твердой уверенности в этом. Количество армян I2* и южноевропейские черты I2a1 M26+, не говоря уже о том, что IJ является родителем как «J», так и «I», склоняют меня к этому».
В I2 есть три клада. Каждый, по-видимому, пошел отдельным путем после последнего ледникового максимума (10 000 лет назад). По словам Кена Нордтведта: «I2 имеет три основных клада, чьи предковые линии разделились 8 000 лет назад или около того. В одном из них есть сильный вклад из Армении/Армянского нагорья и других мест в Юго-Западной Азии. А греческие члены этого клада добавляют ему географической уникальности. У него также есть очень молодой еврейский субклад. Также по словам Кена: «Чем больше я смотрю на общую кладу, которая включает армянский кластер, тем больше я вижу клад, распространенный торговой семьей или кланом по всей широте от Иберии до Кавказа и через Средиземноморье. Где он берет свое начало, определить трудно». Наконец, Кен добавляет: «… два из кладов I2* являются чисто европейскими — и северо-западной и центральной Европой в придачу, хотя включены несколько итальянских гаплотипов. Эти последние два клада I2* имеют оценку возраста межкладового узла в 7500 лет.
Армянские члены клада I2* B имеют возраст коалесценции 2300 лет. При включении еврейских членов I2* того же клада возраст коалесценции становится 3400 лет. Клада A имеет возраст коалесценции 3600 лет в Северной Европе, а клад C имеет возраст коалесценции 2200 лет в Центральной Европе. «Три клада I2* разошлись друг с другом очень давно по сравнению с их индивидуальными возрастами коалесценции. Однажды у нас будут SNP, чтобы различать эти клады». Эта карта текущего распределения различных кладов I2* весьма показательна.
Общий предок семи армянских I2* жил примерно 4500 лет назад на Армянском нагорье.
Вот последний пост на эту тему от Кена Нордтведта: «Кластер I2* является загадкой, потому что в целом I2, вероятно, является самой странной гаплогруппой в Hg I. I2, как правило, распространена по всем четвертям Европы, за исключением заметной слабости в Скандинавии. * имеет сильное и гораздо более древнее армянское присутствие и появляется в других местах на Ближнем Востоке/Леванте. Однако из-за предвзятости наших хороших баз данных объективное утверждение о его относительной силе на этом обширном участке территории пока не может быть сделано. Но ни одна другая подгаплогруппа I не имеет такой географической концентрации. И I2* является старой по времени, возвращаясь к своему TMRCA — только I2a1 M26+ Sardinian оспаривает статус старейшей клады Hg I». Что касается ветви I2a, я все больше склоняюсь к тому, что она представляет собой один из основных генетических компонентов индоевропейского говорящего «армянского» народа, который сам является частью фригийского народа, пришедшего в Анатолию с Запада около 1300 г. до н. э. По словам доктора Роя Кинга: «… ассирийцы и армяне практически идентичны [генетически], за исключением языка, который должен быть отражен в частотах I2 и, возможно, E1b1b1a-V13 для индоевропейского суперстрата. Это интересно тем, что предполагает, что индоевропейские армянские носители пришли с Балкан, а не через Кавказ».
Общий предок семи армянских I2* жил примерно 4500 лет назад на Армянском нагорье.
Здесь вы можете увидеть великолепное ДЕРЕВО гаплогрупп I1 и I2, подготовленное Кеном Нордведтом. В этой версии от мая 2011 года учтены основные открытия и полная переклассификация различных ветвей.
По словам Кена Нордтведта: «Примерно общее разделение гаплогруппы I на четыре ветви 20 000 лет назад, вероятно, имеет некоторые последствия для наших предположений о том, как гаплогруппа I распространилась в Европе (и Анатолии) так давно. Ветвь I2 сейчас еще более древняя».
Гаплогруппа T, ранее известная как K2, имеет две ветви, T1a и T1b, и встречается с низкой частотой по всей Европе и в некоторых частях Ближнего Востока, Северной Африки, как можно увидеть на этой КАРТЕ. Ее происхождение и распространение все еще остаются загадкой, но мы постепенно получаем больше ясности, как можно увидеть на этой новой большой и относительно медленно загружаемой КАРТЕ. Мировое распространение гаплогруппы T неравномерно с некоторыми небольшими областями с концентрацией выше средней. T1a имеет более южный уклон от Ближнего Востока к Европе и Северной Африке. T1b имеет более северный уклон от Ближнего Востока через русские равнины к Северной Европе.
Это мнение Гарета Хенсона, администратора проекта гаплогруппы T в FTDNA: «Я думаю, что и T, и L возникли в регионе Ирака/Ирана, но в то время как ветви L пошли в разных направлениях, я не вижу похожей модели для T. Основные группы — T1a и T1b. Обе, похоже, распространились вместе, но T1a показывает больше вариаций».
В крупной статье о гаплогруппе T, опубликованной в 2011 году, было обнаружено несколько новых мутаций SNP, которые определяют несколько новых ветвей (Fernando L. Mendez et al, “Increased Resolution of Y Chromosome Haplogroup T Defines Relationships among Populations of the Near East, Europe, and Africa”). Общая структура не изменилась, поскольку основными ветвями остаются T1a и T1b. Была обнаружена новая связь с родственной гаплогруппой L.
Участник проекта Жан-Грегуар Манукян подготовил этот увлекательный (но устаревший) синтез гаплогруппы R2. Ранние результаты для армянских участников проекта R2 с 67-маркерными результатами вписываются в это филогенетическое дерево R2 следующим образом. По словам администратора проекта Family Tree DNA R2-WTY, «Возможно, с более разнообразными результатами, которые подтвердят или расширят армянский нагорный кластер, я бы рассмотрел теорию центральноазиатского или анатолийского происхождения гаплогруппы R2 или (по крайней мере, древнего анатолийского кластера). Взгляните на это, чтобы понять, как интерпретировать филогенетическое дерево»
Благодаря глубокому тестированию гаплогруппы R2 начинает вырисовываться структура ветвей. Это структура по состоянию на начало 2011 года на основе недавно выявленных мутаций SNP.
Гаплогруппа L изучена недостаточно. Ее распространение в нашем регионе, по крайней мере, интригует. Есть очаги L в Амшене, Понтийском регионе, ливанских маронитах, живущих на восточной границе Армянской Месопотамии/Ирака, израильских друзах и сирийцах. Структура ветвей этой гаплогруппы не очень хорошо определена. Большинство людей в нашем регионе принадлежат к L2 и ее ветвям. Армяне по-прежнему принадлежат к ветвям L2* и L2a. По словам Гарета Хенсона: «L2 — это ближневосточная и средиземноморская ветвь L, почти полностью отсутствующая в Индии и Пакистане (за исключением, возможно, парсов, которые имеют иранское происхождение, у которых есть отличительный гаплотип L, который я не видел. Были протестированы SNP, но он, похоже, ближе всего к L2).
Следующий HLA был построен на основе опубликованных данных. Отнеситесь к этому с долей скепсиса, поскольку различия в модальном гаплотипе L между Анатолией и Индией предполагают независимые расширения от двух разных популяций-основателей. Нам нужны разные карты для разных ветвей. Надеюсь, скоро будут опубликованы новые научные статьи об этом увлекательном гаплотипе.
Гарет Хенсон. «Я думаю, что и T, и L возникли в регионе Ирака/Ирана… Все ветви L пошли в разных направлениях (L1 на юго-восток, L2 на запад и L3 на северо-восток).
Гаплогруппа Q, возможно, самая молодая из 20 гаплогрупп Y-ДНК, возникла из-за мутации SNP M242+ у человека, который, вероятно, жил в Сибири примерно за 15 000–20 000 лет до настоящего времени. На данном этапе слишком мало известно о субкладах Q1a3 (M346+) и Q1b1a (L245+). Здесь обсуждается одна из теорий происхождения Q1b. Недавно были обнаружены новые SNP. Возможно, они дадут структуру этой ветви. См. проект ДЕРЕВА гаплогруппы Q.
Двое из участников проекта «Армянская ДНК» принадлежат к очень редкой гаплогруппе F, матери всех неафриканских гаплогрупп, за исключением C, D и E, как можно увидеть на этой простой диаграмме и этом более сложном ДЕРЕВЕ.
Человек из гаплогруппы C, F был общим предком всех людей, которые мигрировали за пределы Африки до недавнего времени. Определяющая мутация произошла 31 000–55 000 лет назад на северо-востоке Африки и до сих пор наиболее распространена в Африке в настоящее время — в Эфиопии и Судане.
Армянские F принадлежат к ветви F3, потому что они дали положительный результат (выведенный) на P96 и M282. У одного из них также есть шесть недавно обнаруженных мутаций SNP: L279, L280, L281, L284, L285, L286. Множество новых SNP из секвенирования Family Tree DNA «Walk Through the Y» являются лишь свидетельством длительного времени, прошедшего между нашим армянским «F» и ранее секвенированными мужчинами.
Относительно молодая гаплогруппа R1a (M17+) является наиболее частой гаплогруппой Y-хромосомы среди популяций, представляющих самые разные языковые группы, такие как славянские, индоиранские, дравидийские, которые встречаются в обширной географической области, простирающейся от Южной Азии до Центральной Восточной Европы и Южной Сибири. Пик приходится на Польшу — 35%. Его возраст и распространение показаны на этой КАРТЕ и этой КАРТЕ. Гаплогруппа R1a была обнаружена у 5% армян и в более высоких концентрациях в следующих кавказских группах: 27,5% карачаевцев и балкарцев, абхазов. Ни армяне, ни кавказские группы, похоже, не имеют славянского происхождения, о чем свидетельствует эта КАРТА субклада R1a1a7 (M458+). Нам нужно смотреть на Восток (Иран, Афганистан, Пакистан и Индия) или на Север (Русские равнины) для объяснения присутствия R1a в нашем регионе.
Если у вас есть свободное время, вы можете просмотреть подробные руководства (с устными и визуальными слайдовыми презентациями), подготовленные Genbase по различным семействам гаплогрупп. Поскольку армянская ДНК не была тщательно проанализирована, когда были подготовлены эти руководства, мы не согласны с некоторыми утверждениями относительно происхождения различных кладов и субкладов гаплогрупп:
Гаплогруппа U мтДНК произошла от женщины, которая жила около 55 000 лет назад. Ее потомки дали начало нескольким различным подгруппам, некоторые из которых имеют определенную географическую родину. Древний возраст этой гаплогруппы привел к широкому распространению подгрупп потомков, которые содержат определенные европейские, североафриканские, индийские, арабские, армянские горные и ближневосточные клады. Ветвь U1, по-видимому, появляется в основном на Ближнем Востоке, однако результаты с низкой частотой кажутся разбросанными по всей Европе, особенно в Средиземноморье. U1a встречается от Индии до Европы, но крайне редко встречается среди северных и атлантических окраин Европы. U1b имеет похожее распространение, но встречается реже, чем U1a. Гаплогруппа U2 наиболее распространена в Южной Азии, но также встречается с низкой частотой в Центральной и Западной Азии, а также в Европе как U2e. Эта гаплогруппа была обнаружена в останках 30 000-летнего охотника-собирателя в южноевропейской части России. Ветвь U3, по-видимому, довольно древняя (вероятно, не менее 35 000–45 000 лет). Она встречается с наибольшей частотой в странах, окружающих Черное море, особенно в Болгарии и на Кавказе, а также в некоторых странах Ближнего Востока, таких как Иордания и Сирия, как можно увидеть на этой КАРТЕ. Она встречается в очень низких количествах по всей Европе (около 1% населения), на Ближнем Востоке (около 2,5% населения) и в Центральной Азии (1%). U3 присутствует в высоких количествах среди литовских, польских и испанских цыганских популяций (36–56%). Ветвь U4 берет свое начало в верхнем палеолите, датируемом примерно 25 000 лет назад. Она широко распространена в Европе и была вовлечена в экспансию современных людей в Европу, произошедшую до последнего ледникового максимума. Она встречается с низкой частотой по всей Европе, Северной Америке и Азии. Она особенно распространена в Финляндии и России. Ветвь U5 предшествует расширению сельского хозяйства в Европе. Его возраст оценивается в 50 000 лет, но может достигать 60 500 лет. Гаплогруппа U5 и ее субклады U5a и U5b образуют самые высокие концентрации населения на крайнем севере, у саамов, финнов и эстонцев, но она широко распространена на более низких уровнях по всей Европе. Это распределение и возраст гаплогруппы указывают на то, что люди из этой гаплогруппы были частью первоначальной экспансии, отслеживающей отступление ледяных щитов из Европы ~10 000 лет назад. Гаплогруппа U5 также встречается с небольшими частотами и с гораздо меньшим разнообразием на Ближнем Востоке и в некоторых частях Африки, что предполагает обратную миграцию людей из северной Европы на юг. U5a в шесть раз более распространена, чем U5b в наших регионах, где она достигает 6,3% населения Кавказа и 3,3% населения Ирана. Во многих европейских популяциях отсутствует гаплогруппа U7, но ее частота превышает 4% на Ближнем Востоке и до 5% в Пакистане, достигая почти 10% уровня у иранцев. В Индии частота гаплогруппы U7 достигает пика более 12% в Гуджарате, самом западном штате Индии, в то время как для всей Индии ее частота остается около 2%. Время экспансии и разнообразие гаплотипов для индийских и ближневосточных мтДНК U7 поразительно схожи. Возможная родина этой гаплогруппы охватывает индийский Гуджарат и Иран, потому что оттуда ее частота резко снижается как на восток, так и на запад. Если происхождение было в Иране, а не в Индии, то ее одинаково высокая частота, а также разнообразие в Гуджарате благоприятствуют сценарию, при котором U7 была введена в прибрежную западную Индию либо очень рано, либо несколькими основателями. Вот самое последнее ДЕРЕВО для этой гаплогруппы. А вот полный список последних мутаций U SNP. Армянские участники проекта принадлежат ко всем шести вышеупомянутым субгаплогруппам.
Гаплогруппа H мтДНК, вероятно, развилась в Западной Азии ~30 000 лет назад, а затем прибыла в Европу ~20-25 000 лет назад, быстро распространившись на юго-запад континента. В настоящее время это самая распространенная гаплогруппа мтДНК в Европе, представляющая около половины населения. Эта гаплогруппа также распространена в Северной Африке и на Ближнем Востоке. Субгаплогруппа H13 особенно распространена на Кавказе. Субгаплогруппы H2 и H8 довольно распространены на Кавказе и в Восточной Европе. H5, возможно, развилась в Западной Азии, где она наиболее часто встречается и разнообразна на Кавказе. Вот самое последнее ДЕРЕВО для этой гаплогруппы. Ключевая статья по гаплогруппе H в нашем регионе была опубликована в 2006 году Roostalu: Происхождение и расширение гаплогруппы H, доминирующей линии митохондриальной ДНК человека в Западной Евразии: ближневосточная и кавказская перспектива. Хотя армянские члены этой важной гаплогруппы не были тщательно протестированы, те, кто был, принадлежат к четырем вышеупомянутым подгаплогруппам.
Гаплогруппа W мтДНК встречается в низких количествах от Европы через Ближний Восток и Кавказ вплоть до Индии.
Считается, что гаплогруппа T мтДНК возникла в Месопотамии и/или Армянской Месопотамии (современные Сирия и Турция) более 45 000 лет назад. Основные подгруппы гаплогруппы T появились в Европе около неолитического периода, около 10 000 лет назад. Оказавшись в Европе, эти подгруппы претерпели резкое расширение, связанное с появлением сельского хозяйства. Гаплогруппа T имеет очень широкое распространение и присутствует на востоке до долины Инда, которая граничит с Индией и Пакистаном, и на юге до Аравийского полуострова. Она включает чуть менее 10% современных европейцев. Вот самое последнее ДЕРЕВО этой гаплогруппы.
мтДНК Гаплогруппа I встречается в низких количествах в Европе, на Ближнем Востоке и Армянском нагорье. Считается, что она возникла где-то в Евразии ~30 000 лет назад и была одной из первых гаплогрупп, переместившихся в Европу. Вот самое последнее ДЕРЕВО этой гаплогруппы.
мтДНК Гаплогруппа HV — это западная евразийская гаплогруппа, встречающаяся на всем Ближнем Востоке, включая Иран, Армянское нагорье и Кавказ. Намного раньше, около 30 000 лет назад, некоторые члены HV переместились на север через Кавказские горы и на запад через Анатолию, их родословные впервые были принесены в Европу кроманьонцами. Их прибытие в Европу ознаменовало конец эпохи неандертальцев, гоминидов, населявших Европу и части Западной Азии примерно с 230 000 до 29 000 лет назад. Лучшие навыки общения, оружие и находчивость, вероятно, позволили им превзойти неандертальцев за скудные ресурсы. Важно отметить, что некоторые потомки HV уже отделились и сформировали свою собственную группу, гаплогруппу H, и продолжили продвижение в Западную Европу. Вот самое последнее ДЕРЕВО для этой гаплогруппы.
Гаплогруппа мтДНК X изначально отделилась от гаплогруппы N, а затем еще больше отделилась около 20 000–30 000 лет назад, образовав две подгруппы, X1 и X2. Она составляет около 2% населения Европы, Ближнего Востока и Северной Африки. Подгруппа X1 гораздо менее многочисленна и ограничена Северной и Восточной Африкой, а также Ближним Востоком. Подгруппа X2, по-видимому, претерпела обширное расширение популяции и расселение около или вскоре после последнего ледникового максимума, около 21 000 лет назад. Она сильнее представлена на Ближнем Востоке, Кавказе и в Средиземноморской Европе и несколько слабее представлена в остальной Европе. Особенно высокие концентрации наблюдаются в Грузии (8%), последнее предположительно из-за эффекта основателя. Вот самое последнее ДЕРЕВО для этой гаплогруппы.
mtDNA Гаплогруппа J: Около 45 000 лет назад в ДНК женщины, жившей на Ближнем Востоке или в Армянском нагорье и на Кавказе, произошла мутация. Дальнейшие мутации произошли в линии J, которую можно идентифицировать как J1a1 (27 000 лет назад), J2a (19 000 лет назад), J2b2 (16 000 лет назад), J2b3 (5 800 лет назад) и т. д. Гаплогруппа J (вместе с «T») связана с распространением земледелия и скотоводства в Европе в эпоху неолита (8 000–10 000 лет назад). Все остальные группы западно-евразийского происхождения (H, V, U, K, W, I, X) ранее занимались охотой и собирательством. Средняя частота гаплогруппы J в целом самая высокая на Ближнем Востоке (12%), за которым следуют Европа (11%), Кавказ (8%) и Северная Африка (6%). Из двух основных подгрупп J1 занимает четыре пятых от общего числа и распространена на континенте, в то время как J2 более локализована вокруг Средиземноморья, Греции, Италии/Сардинии и Испании. Вот самое последнее ДЕРЕВО для этой гаплогруппы.
Гаплогруппы мтДНК N1b и N1c особенно распространены на Ближнем Востоке, Ближнем Востоке, Армянском нагорье и Кавказе. Вот самое последнее ДЕРЕВО для этой гаплогруппы. Профессор Маллия Гундер Паланичами изучала hg N1. Недавно он опубликовал статью о N1a, в которой можно найти это ДЕРЕВО. Он планирует вскоре опубликовать статьи о N1b и N1c.
Гаплогруппа мтДНК K относительно молодая. Впервые она появилась около 16 000 лет назад. Сейчас она представляет собой значительную часть генофонда Западной Евразии. В Европе она особенно распространена вокруг Альп и Британских островов. Она встречается реже в Северной Африке, на Ближнем Востоке и в Южной Азии. Концентрации выше среднего у евреев-ашкенази и друзов. Вот самое последнее ДЕРЕВО для этой гаплогруппы.
Считается, что мтДНК Гаплогруппа V возникла примерно 12 000 лет назад, возможно, в Иберии, согласно опубликованной информации. Она происходит от гаплогруппы HV0a, которая также дала начало гаплогруппе H. За исключением редких, изолированных случаев, V практически отсутствует на Южных Балканах, Армянском нагорье, Кавказе и Ближнем Востоке. Подтвержденных миграций с севера (запада) в эти регионы в исторические времена, безусловно, достаточно, чтобы объяснить эти несколько «беспорядочных» ветвей. У меня есть ощущение, что предыдущее объяснение может не сработать, поскольку теперь мы знаем о трех армянских V. Только время покажет, откуда возникла эта группа. Ключевая статья по гаплогруппе V i была опубликована в 2001 году Торрони и др. Теперь он довольно устарел: Сигнал, от человеческой мтДНК, о постледниковой реколонизации в Европе Вот самое последнее ДЕРЕВО для этой гаплогруппы.
мтДНК Гаплогруппа F сосредоточена в Азии и появляется в восточном Китае и Японии. Она не обнаружена в Америке. Образцы F1b1 были обнаружены в Армении, Турции, Мазандаране, на Кавказе, в Грузии и Хорватии. Это потомок гаплогруппы гаплогруппы R. Вот самое последнее ДЕРЕВО.
Гаплогруппа мтДНК M1 — единственный вариант M, обнаруженный в Африке. Ограниченное распространение гаплогруппы M в Африке и ее широкое распространение за пределами Африки позволяют предположить, что эта линия возникла очень близко ко времени миграции из Африки, либо незадолго до, либо вскоре после выхода из Африки. M1 относительно распространена в Средиземноморье, достигнув пика в Иберии. Она также широко распространена на Ближнем Востоке, от юга Аравийского полуострова до Анатолии и от Леванта до Ирана. Кроме того, гаплотипы M1 иногда наблюдались на Кавказе и в Армянском нагорье. Вот самое последнее ДЕРЕВО для этой гаплогруппы.
Если вы сравниваете свою последовательность мтДНК с другой последовательностью из той же ветви гаплогруппы, обратите внимание на следующее: разницу в одну мутацию в HVR1 или HVR2 следует принимать за 400 лет, а разницу в одну мутацию в области кодирования CR следует принимать за 1600 лет.
R1b1a1
R1b1a1 (название 2011 года) определяется наличием маркера SNP M73. Он был обнаружен с низкой частотой в целом по всей Центральной Евразии, но был обнаружен с относительно высокой частотой среди определенных популяций там, включая хазарейцев в Пакистане (8/25 = 32%);. Четыре мужчины R-M73 были также обнаружены в исследовании 523 человек в западной части Армянского нагорья, и один человек в исследовании 168 человек на Крите.
В 2010 году Майрес и др. сообщают, что из 193 мужчин R-M73, обнаруженных среди 10 355 широко распространенных мужчин, «все, за исключением двух русских, произошли за пределами Европы, либо на Кавказе, либо в западной части Армянского нагорья, либо в регионах Северного Пакистана».

You must be logged in to post a comment.